Треба скласти одне речення зі слів: Узбережжя, згадувати, морський, мріяти, туди, знову, потрапити, насолоджуватися, щоб, краса. Інші слова не можна вставляти …
Ответы (2)
допоможіть, дуже прошу випишіть словосполучення, визначте їх вид (узгодження, керування, прилягання) Хліб і сіль, черговий уряд, утричі швидше, перша спроба, далеко від дому, виконуючи завдання, мороз і сонце, найбільш стійкий
Ответы (1)
Помогите подобрать: Хтось (з, зі,із) нас. Дорога (з,із, зі) міста. тесляр (з,із, зі) хутора. Олена (з,із, зі) матер’ю. розмовляла (з,із, зі) батьком. онука (з,із, зі) дідусем. Оглядачь (з, зі,із) Львова. зустрічався з артистами.
Ответы (2)
| В Википедии есть статья «мегалодон». |
| В Викиданных есть лексема мегалодон (L126178). |
Морфологические и синтаксические свойства[править]
| падеж | ед. ч. | мн. ч. |
|---|---|---|
| Им. | мегалодо́н | мегалодо́ны |
| Р. | мегалодо́на | мегалодо́нов |
| Д. | мегалодо́ну | мегалодо́нам |
| В. | мегалодо́на | мегалодо́нов |
| Тв. | мегалодо́ном | мегалодо́нами |
| Пр. | мегалодо́не | мегалодо́нах |
ме—га—ло—до́н
Существительное, одушевлённое, мужской род, 2-е склонение (тип склонения 1a по классификации А. А. Зализняка).
Корень: —.
Произношение[править]
- МФА: ед. ч. [mʲɪɡəɫɐˈdon], мн. ч. [mʲɪɡəɫɐˈdonɨ]
Семантические свойства[править]
Значение[править]
- ихтиол., палеонт. вымершая акула из семейства Otodontidae, существовавшая в миоцене и плиоцене, одна из самых больших рыб всех времён ◆ Отсутствует пример употребления (см. рекомендации).
Синонимы[править]
- —
Антонимы[править]
- —
Гиперонимы[править]
- акула, рыба
Гипонимы[править]
- —
Родственные слова[править]
| Ближайшее родство | |
Этимология[править]
От ??
Фразеологизмы и устойчивые сочетания[править]
Перевод[править]
| Список переводов | |
Библиография[править]
|
|
Для улучшения этой статьи желательно:
|
Українська мова | 1 — 4 классы
Как пишется по английский мегалодон.

Ninahard
23 нояб. 2019 г., 01:13:38 | 1 — 4 классы
Как пишеться «Духм?
Как пишеться «Духм.
Яний.
На странице вопроса Как пишется по английский мегалодон? из категории Українська мова вы найдете
ответ для уровня учащихся 1 — 4 классов. Если полученный ответ не
устраивает и нужно расшить круг поиска, используйте удобную поисковую
систему сайта. Можно также ознакомиться с похожими вопросами и ответами
других пользователей в этой же категории или создать новый вопрос. Возможно,
вам будет полезной информация, оставленная пользователями в комментариях, где
можно обсудить тему с помощью обратной связи.
Последние ответы

0506691143
4 мая 2022 г., 11:41:15
Обійшов — дієслово, поч.
Ф. — обійти, доконаний вид, перехідне, І дієвідміна, дійсний спосіб, минулий час, ІІІ особа, однина, чоловічий рід, присудок.
Присудок — це скоріше за все, бо я не знаю речення…
Слово Обійшов, зробити розбір як частини мови?

Yuliaqx
4 мая 2022 г., 10:50:13
Народився на Полтавщині.
Навчався в Полтавській духовній семінарії, та покинув цей духовний заклад і перейшов до Харківського ветеринарного інституту.
Закінчивши ВУЗ у 1902 році, працював ветеринарним лікарем.
Видав популярний посібник з ветеринар..
Відомості про василя короліва старого?

GR1F
4 мая 2022 г., 10:20:58
Нас оточує багата природа!
Гори, ріки, ліси — це все багатства природи.
Все це дуже красиве, але є люди які цього не розуміють і просто нищуть ці багатства.
Уявіть собі скільки людей на землі і якщо люди не будуть дбати про природу її не буде.
Ос..
Помогите пожалуйста текс на тему , , Бережить природу»?

КАРЯК
4 мая 2022 г., 09:01:38
Обереги є давніми, добрими символами нашого народу.
До оберегів належать вишиті рушники, писанки, вишиванки, підкови, вінок, вербові гілки, маківки, образи.
З давних давен вважали , що вишиванка захищає від усілякого лиха, наділяє людину цілющою си..
Твір роздум на тему родинні обереги?

0КиСуЛя0
4 мая 2022 г., 08:22:53
Синонимы — белый, прозрачный
Антонимы — грязный, чёрный
Общеупотребл.
Слово
Фразеологизмы — чистой воды, принять за чистоту!
)))…
Лексичний анализ слова чысті?

3993
4 мая 2022 г., 06:42:47
Легенда — закiнчення а, корiнь легенд.
Монастир — закiнчення нульове, корiнь — монастир.
Горi — закiнчення i, корiнь гор.
Джерельця — закiнчення я, корiнь джерель, суфiкс ц
Неба — закiнчення а, корiнь неб
Слiдочок — закiнчення ок, корiнь слiд, суф..
Позначити у словах корінь, префікс, суфікс : легенда, монастир, горі, джерельця, неба, слідочок, пре
Слово мегалодон имеет такое значение:
— вымерший вид акул;
| Megalodon
Temporal range: Early Miocene–Early Pliocene, c. 23–3.6 Ma PreꞒ Ꞓ O S D C P T J K Pg N |
|
|---|---|

|
|
| Model of megalodon jaws at the American Museum of Natural History | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Otodontidae |
| Genus: | †Otodus |
| Species: |
†O. megalodon |
| Binomial name | |
| †Otodus megalodon
(Agassiz, 1843)[1] |
|
| Synonyms[2][3][4][5][6] | |
|
List of synonyms
|
Megalodon (Otodus megalodon),[6][7][8] meaning «big tooth», is an extinct species of mackerel shark that lived approximately 23 to 3.6 million years ago (Mya), from the Early Miocene to the Pliocene epochs.[9] It was formerly thought to be a member of the family Lamnidae and a close relative of the great white shark (Carcharodon carcharias). However, it is now classified into the extinct family Otodontidae, which diverged from the great white shark during the Early Cretaceous.
While regarded as one of the largest and most powerful predators to have ever lived, the megalodon is only known from fragmentary remains, and its appearance and maximum size are uncertain. Scientists differ on whether it would have more closely resembled a stockier version of the great white shark, the whale shark (Rhincodon typus), the basking shark (Cetorhinus maximus) or the sand tiger shark (Carcharias taurus). The most recent estimate with the least error range suggests a maximum length estimate up to 20.3 meters (67 ft),[10] although the modal lengths are estimated at 10.5 meters (34 ft).[11][12] Extrapolation from a vertebral centra with dimensions based on the great white shark suggests that a megalodon about 16 meters (52 ft) long weighs up to 48 metric tons (53 short tons), 17 meters (56 ft) long weighs up to 59 metric tons (65 short tons), and 20.3 meters (67 ft) long (the maximum length) weighs up to 103 metric tons (114 short tons).[13][14] Extrapolating from a vertebral column and reconstructing a 3D model with dimensions based on all extant lamnid sharks suggests that a 16-meter-long (52 ft) individual may have been much larger than previous estimates, reaching an excess of 61.5 metric tons (67.8 short tons) in body mass; an individual of this size would have needed to consume 98,175 kcal per day.[15] Their teeth were thick and robust, built for grabbing prey and breaking bone, and their large jaws could exert a bite force of up to 108,500 to 182,200 newtons (24,400 to 41,000 lbf).[14]
Megalodon probably had a major impact on the structure of marine communities. The fossil record indicates that it had a cosmopolitan distribution. It probably targeted large prey, such as whales, seals and sea turtles. Juveniles inhabited warm coastal waters and fed on fish and small whales. Unlike the great white, which attacks prey from the soft underside, megalodon probably used its strong jaws to break through the chest cavity and puncture the heart and lungs of its prey.
The animal faced competition from whale-eating cetaceans, such as Livyatan and other macroraptorial sperm whales and possibly smaller ancestral killer whales. As the shark preferred warmer waters, it is thought that oceanic cooling associated with the onset of the ice ages, coupled with the lowering of sea levels and resulting loss of suitable nursery areas, may have also contributed to its decline. A reduction in the diversity of baleen whales and a shift in their distribution toward polar regions may have reduced megalodon’s primary food source. The shark’s extinction coincides with a gigantism trend in baleen whales.
Taxonomy
Naming
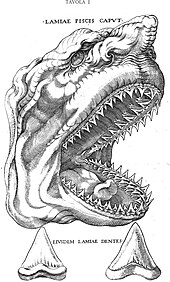
The depiction of a shark’s head by Nicolas Steno in his work The Head of a Shark Dissected
According to Renaissance accounts, gigantic triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolas Steno, who recognized them as shark teeth, and famously produced a depiction of a shark’s head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a megalodon tooth.[16][17][18]
Swiss naturalist Louis Agassiz gave this shark its initial scientific name, Carcharodon megalodon, in his 1843 work Recherches sur les poissons fossiles, based on tooth remains.[1][19] English paleontologist Edward Charlesworth in his 1837 paper used the name Carcharias megalodon, while citing Agassiz as the author, indicating that Agassiz described the species prior to 1843. English paleontologist Charles Davies Sherborn in 1928 listed an 1835 series of articles by Agassiz as the first scientific description of the shark.[20] The specific name megalodon translates to «big tooth», from Ancient Greek: μέγας, romanized: (mégas), lit. ‘big, mighty’ and ὀδούς (odoús), «tooth».[21][22] The teeth of megalodon are morphologically similar to those of the great white shark (Carcharodon carcharias), and on the basis of this observation, Agassiz assigned megalodon to the genus Carcharodon.[19]
There was one apparent description of the shark in 1881 classifying it as Selache manzonii.[23]
Evolution

While the earliest megalodon remains have been reported from the Late Oligocene, around 28 million years ago (Mya),[24][25] there is disagreement as to when it appeared, with dates ranging to as young as 16 mya.[26] It has been thought that megalodon became extinct around the end of the Pliocene, about 2.6 Mya;[26][27] claims of Pleistocene megalodon teeth, younger than 2.6 million years old, are considered unreliable.[27] A 2019 assessment moves the extinction date back to earlier in the Pliocene, 3.6 Mya.[28]
Megalodon is now considered to be a member of the family Otodontidae, genus Otodus, as opposed to its previous classification into Lamnidae, genus Carcharodon.[26][12][27][6][7] Megalodon’s classification into Carcharodon was due to dental similarity with the great white shark, but most authors currently believe that this is due to convergent evolution. In this model, the great white shark is more closely related to the extinct broad-toothed mako (Isurus hastalis) than to megalodon, as evidenced by more similar dentition in those two sharks; megalodon teeth have much finer serrations than great white shark teeth. The great white shark is more closely related to the mako shark (Isurus spp.), with a common ancestor around 4 Mya.[19][29] Proponents of the former model, wherein megalodon and the great white shark are more closely related, argue that the differences between their dentition are minute and obscure.[30]: 23–25

The genus Carcharocles currently contains four species: C. auriculatus, C. angustidens, C. chubutensis, and C. megalodon.[31]: 30–31 The evolution of this lineage is characterized by the increase of serrations, the widening of the crown, the development of a more triangular shape, and the disappearance of the lateral cusps.[31]: 28–31 [32] The evolution in tooth morphology reflects a shift in predation tactics from a tearing-grasping bite to a cutting bite, likely reflecting a shift in prey choice from fish to cetaceans.[33] Lateral cusplets were finally lost in a gradual process that took roughly 12 million years during the transition between C. chubutensis and C. megalodon.[33] The genus was proposed by D. S. Jordan and H. Hannibal in 1923 to contain C. auriculatus. In the 1980s, megalodon was assigned to Carcharocles.[19][31]: 30 Before this, in 1960, the genus Procarcharodon was erected by French ichthyologist Edgard Casier, which included those four sharks and was considered separate from the great white shark. It is now considered a junior synonym of Carcharocles.[31]: 30 The genus Palaeocarcharodon was erected alongside Procarcharodon to represent the beginning of the lineage, and, in the model wherein megalodon and the great white shark are closely related, their last common ancestor. It is believed to be an evolutionary dead-end and unrelated to the Carcharocles sharks by authors who reject that model.[30]: 70

The great white shark (Carcharodon carcharias) and megalodon were previously thought to be close relatives.[19][29]
Another model of the evolution of this genus, also proposed by Casier in 1960, is that the direct ancestor of the Carcharocles is the shark Otodus obliquus, which lived from the Paleocene through the Miocene epochs, 60 to 13 Mya.[29][32] The genus Otodus is ultimately derived from Cretolamna, a shark from the Cretaceous period.[6][34] In this model, O. obliquus evolved into O. aksuaticus, which evolved into C. auriculatus, and then into C. angustidens, and then into C. chubutensis, and then finally into C. megalodon.
Another model of the evolution of Carcharocles, proposed in 2001 by paleontologist Michael Benton, is that the three other species are actually a single species of shark that gradually changed over time between the Paleocene and the Pliocene, making it a chronospecies.[31]: 17 [25][35] Some authors suggest that C. auriculatus, C. angustidens, and C. chubutensis should be classified as a single species in the genus Otodus, leaving C. megalodon the sole member of Carcharocles.[25][36]
The genus Carcharocles may be invalid, and the shark may actually belong in the genus Otodus, making it Otodus megalodon.[4] A 1974 study on Paleogene sharks by Henri Cappetta erected the subgenus Megaselachus, classifying the shark as Otodus (Megaselachus) megalodon, along with O. (M.) chubutensis. A 2006 review of Chondrichthyes elevated Megaselachus to genus, and classified the sharks as Megaselachus megalodon and M. chubutensis.[4] The discovery of fossils assigned to the genus Megalolamna in 2016 led to a re-evaluation of Otodus, which concluded that it is paraphyletic, that is, it consists of a last common ancestor but it does not include all of its descendants. The inclusion of the Carcharocles sharks in Otodus would make it monophyletic, with the sister clade being Megalolamna.[6]
The cladogram below represents the hypothetical relationships between megalodon and other sharks, including the great white shark. Modified from Shimada et al. (2016),[6] Ehret et al., (2009),[29] and the findings of Siversson et al. (2013).[38][39]
Biology
Appearance

One interpretation on how megalodon appeared was that it was a robust-looking shark, and may have had a similar build to the great white shark. The jaws may have been blunter and wider than the great white, and the fins would have also been similar in shape, though thicker due to its size. It may have had a pig-eyed appearance, in that it had small, deep-set eyes.[40]
Another interpretation is that megalodon bore a similarity to the whale shark (Rhincodon typus) or the basking shark (Cetorhinus maximus). The tail fin would have been crescent-shaped, the anal fin and second dorsal fin would have been small, and there would have been a caudal keel present on either side of the tail fin (on the caudal peduncle). This build is common in other large aquatic animals, such as whales, tuna, and other sharks, in order to reduce drag while swimming. The head shape can vary between species as most of the drag-reducing adaptations are toward the tail-end of the animal.[31]: 35–36

Sculpture in the Museum of Evolution in Puebla, Mexico
Since Carcharocles is derived from Otodus, and the two had teeth that bear a close similarity to those of the sand tiger shark (Carcharias taurus), megalodon may have had a build more similar to the sand tiger shark than to other sharks. This is unlikely since the sand tiger shark is a carangiform swimmer which requires faster movement of the tail for propulsion through the water than the great white shark, a thunniform swimmer.[31]: 35–36 [41]
Size
Due to fragmentary remains, there have been many contradictory size estimates for megalodon, as they can only be drawn from fossil teeth and vertebrae.[42]: 87 [43] The great white shark has been the basis of reconstruction and size estimation, as it is regarded as the best analogue to megalodon. Several total length estimation methods have been produced from comparing megalodon teeth and vertebrae to those of the great white.[40][44][10][7]

Size comparison of the great white and whale shark to estimates for megalodon

Proportions of megalodon at lengths of 3 m (10 ft), 8 m (26 ft), and 16 m (52 ft), extrapolated from extant relatives, with a 1.65 m (5 ft 5 in) diver
Megalodon size estimates vary depending on the method used, with maximum total length estimates ranging from 14.2–20.3 meters (47–67 ft).[40][10][7] A 2015 study estimated the average total body length at 10.5 meters (34 ft), calculated from 544 megalodon teeth, found throughout geological time and geography, including adults and juveniles.[11][12] In comparison, large great white sharks are generally around 6 meters (20 ft) in length, with a few contentious reports suggesting larger sizes.[45][46][40] The whale shark is the largest living fish, with one large female reported with a precaudal length of 15 meters (49 ft) and an estimated total length of 18.8 meters (62 ft).[45][47] It is possible that different populations of megalodon around the globe had different body sizes and behaviors due to different ecological pressures.[12] Megalodon is thought to have been the largest macropredatory shark that ever lived.[40]
«A C. megalodon about 16 meters long would have weighed about 48 metric tons (53 tons). A 17-meter (56-foot) C. megalodon would have weighed about 59 metric tons (65 tons), and a 20.3-meter (67 foot) monster would have topped off at 103 metric tons (114 tons).»[13]
In his 2015 book, The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution, Donald Prothero proposed the body mass estimates for different individuals of different length by extrapolating from a vertebral centra based on the dimensions of the great white,[13] a methodology also used for the 2008 study which supports the maximum mass estimate.[14]
In 2020, Cooper and his colleagues reconstructed a 2D model of megalodon based on the dimensions of all the extant lamnid sharks and suggested that a 16 meters (52 ft) long megalodon would have had a 4.65 m (15.3 ft) long head, 1.41 m (4 ft 8 in) tall gill slits, a 1.62 m (5 ft 4 in) tall dorsal fin, 3.08 m (10 ft 1 in) long pectoral fins, and a 3.85 m (12 ft 8 in) tall tail fin.[8] In 2022, Cooper and his colleagues also reconstructed a 3D model with the same basis as the 2020 study, resulting in a body mass estimate of 61.56 metric tons (67.86 short tons) for a 16 meters (52 ft) long megalodon (higher than the previous estimates); a vertebral column specimen named IRSNB P 9893 (formerly IRSNB 3121), belonging to a 46 year old individual from Belgium, was used for extrapolation. An individual of this size would have required 98,175 kcal per day, 20 times more than what the adult great white requires.[15]
Mature male megalodon may have had a body mass of 12.6 to 33.9 metric tons (13.9 to 37.4 short tons), and mature females may have been 27.4 to 59.4 metric tons (30.2 to 65.5 short tons), assuming that males could range in length from 10.5 to 14.3 meters (34 to 47 ft) and females 13.3 to 17 meters (44 to 56 ft).[40]
A 2015 study linking shark size and typical swimming speed estimated that megalodon would have typically swum at 18 kilometers per hour (11 mph)–assuming that its body mass was typically 48 metric tons (53 short tons)–which is consistent with other aquatic creatures of its size, such as the fin whale (Balaenoptera physalus) which typically cruises at speeds of 14.5 to 21.5 km/h (9.0 to 13.4 mph).[48] In 2022, Cooper and his colleagues converted this calculation into relative cruising speed (body lengths per second), resulting in an mean absolute cruising speed of 5 kilometers per hour (3.1 mph) and a mean relative cruising speed of 0.09 body lengths per second for a 16 meters (52 ft) long megalodon; the authors found their mean absolute cruising speed to be faster than any extant lamnid sharks and their mean relative cruising speed to be slower, consistent with previous estiamtes.[15]
Its large size may have been due to climatic factors and the abundance of large prey items, and it may have also been influenced by the evolution of regional endothermy (mesothermy) which would have increased its metabolic rate and swimming speed. The otodontid sharks have been considered to have been ectotherms, so on that basis megalodon would have been ectothermic. However, the largest contemporary ectothermic sharks, such as the whale shark, are filter feeders, while lamnids are now known to be regional endotherms, implying some metabolic correlations with a predatory lifestyle. These considerations, as well as tooth oxygen isotopic data and the need for higher burst swimming speeds in macropredators of endothermic prey than ectothermy would allow, imply that otodontids, including megalodon, were probably regional endotherms.[49]
In 2020, Shimada and colleagues suggested large size was instead due to intrauterine cannibalism, where the larger fetus eats the smaller fetus, resulting in progressively larger and larger fetuses, requiring the mother to attain even greater size as well as caloric requirements which would have promoted endothermy. Males would have needed to keep up with female size in order to still effectively copulate (which probably involved latching onto the female with claspers, like modern cartilaginous fish).[50]
Maximum estimates
The first attempt to reconstruct the jaw of megalodon was made by Bashford Dean in 1909, displayed at the American Museum of Natural History. From the dimensions of this jaw reconstruction, it was hypothesized that megalodon could have approached 30 meters (98 ft) in length. Dean had overestimated the size of the cartilage on both jaws, causing it to be too tall.[51][52]
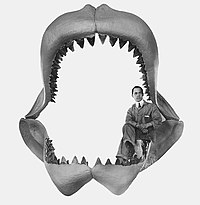

Tooth compared to hand
In 1973, John E. Randall, an ichthyologist, used the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) to measure the length of the shark, yielding a maximum length of about 13 meters (43 ft).[53] However, tooth enamel height does not necessarily increase in proportion to the animal’s total length.[30]: 99
In 1994, marine biologists Patrick J. Schembri and Stephen Papson opined that O. megalodon may have approached a maximum of around 24 to 25 meters (79 to 82 ft) in total length.[54][55]
In 1996, shark researchers Michael D. Gottfried, Leonard Compagno, and S. Curtis Bowman proposed a linear relationship between the great white shark’s total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in meters = − (0.096) × [UA maximum height (mm)]-(0.22).[56][40] Using this tooth height regression equation, the authors estimated a total length of 15.9 meters (52 ft) based on a tooth 16.8 centimeters (6.6 in) tall, which the authors considered a conservative maximum estimate. They also compared the ratio between the tooth height and total length of large female great whites to the largest megalodon tooth. A 6-meter (20 ft) long female great white, which the authors considered the largest ‘reasonably trustworthy’ total length, produced an estimate of 16.8 meters (55 ft). However, based on the largest female great white reported, at 7.1 meters (23 ft), they estimated a maximum estimate of 20.2 meters (66 ft).[40]
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimeter (0.39 in) of root width, there are approximately 1.4 meters (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah’s possession had a root width of about 12 centimeters (4.7 in), which yielded 16.5 meters (54 ft) in total length.[31]: 88
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length after conducting anatomical analysis of several specimens, allowing any sized tooth to be used. Shimada stated that the previously proposed methods were based on a less-reliable evaluation of the dental homology between megalodon and the great white shark, and that the growth rate between the crown and root is not isometric, which he considered in his model. Using this model, the upper anterior tooth possessed by Gottfried and colleagues corresponded to a total length of 15 meters (49 ft).[57] Among several specimens found in the Gatún Formation of Panama, one upper lateral tooth was used by other researchers to obtain a total length estimate of 17.9 meters (59 ft) using this method.[36][58]
In 2019, Shimada revisited the size of megalodon and discouraged using non-anterior teeth for estimations, noting that the exact position of isolated non-anterior teeth is difficult to identify. Shimada provided maximum total length estimates using the largest anterior teeth available in museums. The tooth with the tallest crown height known to Shimada, NSM PV-19896, produced a total length estimate of 14.2 meters (47 ft). The tooth with the tallest total height, FMNH PF 11306, was reported at 16.8 centimeters (6.6 in). However, Shimada remeasured the tooth and found it actually to measure 16.2 centimeters (6.4 in). Using the total height tooth regression equation proposed by Gottfried and colleagues produced an estimate of 15.3 meters (50 ft).[7][10]
In 2021, Victor J. Perez, Ronny M. Leder, and Teddy Badaut proposed a method of estimating total length of megalodon from the sum of the tooth crown widths. Using more complete megalodon dentitions, they reconstructed the dental formula and then made comparisons to living sharks. The researchers noted that the 2002 Shimada crown height equations produce wildly varying results for different teeth belonging to the same shark (range of error of ± 9 metres (30 ft)), casting doubt on some of the conclusions of previous studies using that method. Using the largest tooth available to the authors, GHC 6, with a crown width of 13.3 centimeters (5.2 in), they estimated a maximum body length of approximately 20 meters (66 ft), with a range of error of approximately ± 3.5 metres (11 ft).[10] This maximum length estimate was also supported by Cooper and his colleagues in 2022.[15]
There are anecdotal reports of teeth larger than those found in museum collections.[7] Gordon Hubbell from Gainesville, Florida, possesses an upper anterior megalodon tooth whose maximum height is 18.4 centimeters (7.25 in), one of the largest known tooth specimens from the shark.[59] In addition, a 2.7-by-3.4-meter (9 by 11 ft) megalodon jaw reconstruction developed by fossil hunter Vito Bertucci contains a tooth whose maximum height is reportedly over 18 centimeters (7 in).[60]
Teeth and bite force

The most common fossils of megalodon are its teeth. Diagnostic characteristics include a triangular shape, robust structure, large size, fine serrations, a lack of lateral denticles, and a visible V-shaped neck (where the root meets the crown).[30]: 55 [36] The tooth met the jaw at a steep angle, similar to the great white shark. The tooth was anchored by connective tissue fibers, and the roughness of the base may have added to mechanical strength.[61] The lingual side of the tooth, the part facing the tongue, was convex; and the labial side, the other side of the tooth, was slightly convex or flat. The anterior teeth were almost perpendicular to the jaw and symmetrical, whereas the posterior teeth were slanted and asymmetrical.[62]
Megalodon teeth can measure over 180 millimeters (7.1 in) in slant height (diagonal length) and are the largest of any known shark species,[31]: 33 implying it was the largest of all macropredatory sharks.[40] In 1989, a nearly complete set of megalodon teeth was discovered in Saitama, Japan. Another nearly complete associated megalodon dentition was excavated from the Yorktown Formations in the United States, and served as the basis of a jaw reconstruction of megalodon at the National Museum of Natural History (USNM). Based on these discoveries, an artificial dental formula was put together for megalodon in 1996.[30]: 55 [63]
The dental formula of megalodon is: 2.1.7.43.0.8.4. As evident from the formula, megalodon had four kinds of teeth in its jaws: anterior, intermediate, lateral, and posterior. Megalodon’s intermediate tooth technically appears to be an upper anterior and is termed as «A3» because it is fairly symmetrical and does not point mesially (side of the tooth toward the midline of the jaws where the left and right jaws meet). Megalodon had a very robust dentition,[30]: 20–21 and had over 250 teeth in its jaws, spanning 5 rows.[31]: iv It is possible that large megalodon individuals had jaws spanning roughly 2 meters (6.6 ft) across.[31]: 129 The teeth were also serrated, which would have improved efficiency in cutting through flesh or bone.[19][31]: 1 The shark may have been able to open its mouth to a 75° angle, though a reconstruction at the USNM approximates a 100° angle.[40]

In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5-meter (8.2 ft) long specimen, and then isometrically scaled the results for its maximum size and the conservative minimum and maximum body mass of megalodon. They placed the bite force of the latter between 108,514 to 182,201 newtons (24,395 to 40,960 lbf) in a posterior bite, compared to the 18,216 newtons (4,095 lbf) bite force for the largest confirmed great white shark, and 7,400 newtons (1,700 lbf) for the placoderm fish Dunkleosteus. In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the force generated, which would probably have caused the total force experienced by prey to be higher than the estimate.[14][64]
In 2021, Antonio Ballell and Humberto Ferrón used Finite Element Analysis modeling to examine the stress distribution of three types of megalodon teeth and closely related mega-toothed species when exposed to anterior and lateral forces, the latter of which would be generated when a shark shakes its head to tear through flesh. The resulting simulations identified higher levels of stress in megalodon teeth under lateral force loads compared to its precursor species such as O. obliquus and O. angusteidens when tooth size was removed as a factor. This suggests that megalodon teeth were of a different functional significance than previously expected, challenging prior interpretations that megalodon’s dental morphology was primarily driven by a dietary shift towards marine mammals. Instead, the authors proposed that it was a byproduct of an increase in body size caused by heterochronic selection.[65]
Internal anatomy

Megalodon is represented in the fossil record by teeth, vertebral centra, and coprolites.[40][66] As with all sharks, the skeleton of megalodon was formed of cartilage rather than bone; consequently most fossil specimens are poorly preserved.[67] To support its large dentition, the jaws of megalodon would have been more massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. Its chondrocranium, the cartilaginous skull, would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size.[40]
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp Basin, Belgium, in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimeters (2.2 in) to 155 millimeters (6 in) in diameter. The shark’s vertebrae may have gotten much bigger, and scrutiny of the specimen revealed that it had a higher vertebral count than specimens of any known shark, possibly over 200 centra; only the great white approached it.[40] Another partially preserved vertebral column of a megalodon was excavated from the Gram Formation in Denmark in 1983, which comprises 20 vertebral centra, with the centra ranging from 100 millimeters (4 in) to 230 millimeters (9 in) in diameter.[61]

The coprolite remains of megalodon are spiral-shaped, indicating that the shark may have had a spiral valve, a corkscrew-shaped portion of the lower intestines, similar to extant lamniform sharks. Miocene coprolite remains were discovered in Beaufort County, South Carolina, with one measuring 14 cm (5.5 in).[66]
Gottfried and colleagues reconstructed the entire skeleton of megalodon, which was later put on display at the Calvert Marine Museum in the United States and the Iziko South African Museum.[40][32] This reconstruction is 11.3 meters (37 ft) long and represents a mature male,[40]: 61 based on the ontogenetic changes a great white shark experiences over the course of its life.[40]: 65
Paleobiology
Range and habitat
Megalodon had a cosmopolitan distribution;[26][58] its fossils have been excavated from many parts of the world, including Europe, Africa, the Americas, and Australia.[30]: 67 [68] It most commonly occurred in subtropical to temperate latitudes.[26][30]: 78 It has been found at latitudes up to 55° N; its inferred tolerated temperature range was 1–24 °C (34–75 °F).[note 1] It arguably had the capacity to endure such low temperatures due to mesothermy, the physiological capability of large sharks to maintain a higher body temperature than the surrounding water by conserving metabolic heat.[26]
Megalodon inhabited a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult megalodon were not abundant in shallow water environments, and mostly inhabited offshore areas. Megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.[31]: 33 [70]
Fossil remains show a trend for specimens to be larger on average in the Southern Hemisphere than in the Northern, with mean lengths of 11.6 and 9.6 meters (38 and 31 ft), respectively; and also larger in the Pacific than the Atlantic, with mean lengths of 10.9 and 9.5 meters (36 and 31 ft) respectively. They do not suggest any trend of changing body size with absolute latitude, or of change in size over time (although the Carcharocles lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 meters (34 ft), with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.[12]
Locations of fossils
Megalodon had a global distribution and fossils of the shark have been found in many places around the world, bordering all oceans of the Neogene.[71]
| Epoch | Formation | State | Continent |
|---|---|---|---|
| Pliocene | Luanda Formation | Africa | |
| Africa | |||
| Africa | |||
| Castell’Arquato Formation | Europe | ||
| Arenas de Huelva Formation | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Touril Complex Formation | Europe | ||
| Red Crag Formation | Europe | ||
| San Mateo Formation | North America | ||
| Towsley Formation | North America | ||
| Bone Valley Formation | North America | ||
| Tamiami Formation | North America | ||
| Yorktown Formation | North America | ||
| Highlands Formation | North America | ||
| Refugio Formation | North America | ||
| San Diego Formation | North America | ||
| Tirabuzon Formation | North America | ||
| Onzole Formation | South America | ||
| Paraguaná Formation | South America | ||
| Black Rock Sandstone | Oceania | ||
| Cameron Inlet Formation | Oceania | ||
| Grange Burn Formation | Oceania | ||
| Loxton Sand Formation | Oceania | ||
| Whaler’s Bluff Formation | Oceania | ||
| Tangahoe Formation | Oceania | ||
| Miocene | |||
| Africa | |||
| Madagascar Basin | Africa | ||
| Africa | |||
| Varswater Formation | Africa | ||
| Baripada Limestone | Asia | ||
| Arakida Formation | Asia | ||
| Bihoku Group | Asia | ||
| Fujina Formation | Asia | ||
| Hannoura Formation | Asia | ||
| Hongo Formation | Asia | ||
| Horimatsu Formation | Asia | ||
| Ichishi Formation | Asia | ||
| Kurahara Formation | Asia | ||
| Maenami Formation | Asia | ||
| Matsuyama Group | Asia | ||
| Sekinobana Formation | Asia | ||
| Suso Formation | Asia | ||
| Takakubo Formation | Asia | ||
| Tonokita Formation | Asia | ||
| Tsurushi Formation | Asia | ||
| Wajimazaki Formation | Asia | ||
| Yoshii Formation | Asia | ||
| Asia | |||
| Burgeschleinitz Formation | Europe | ||
| Melker Sand Formation | Europe | ||
| Rzehakia Formation | Europe | ||
| Weissenegg Formation | Europe | ||
| Antwerpen Sands Member | Europe | ||
| Europe | |||
| Hrušky Formation | Europe | ||
| Gram Formation | Europe | ||
| Aquitaine Basin | Europe | ||
| Europe | |||
| Libano Sandstone | Europe | ||
| Blue Clay Formation | Europe | ||
| Globigerina Limestone[72] | Europe | ||
| Aalten Member | Europe | ||
| Breda Formation | Europe | ||
| Korytnica Clays | Europe | ||
| Leitha Limestone | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Filakovo Formation | Europe | ||
| Arjona Formation | Europe | ||
| Calcarenites of Sant Elm | Europe | ||
| Europe | |||
| Monterey Formation | North America | ||
| Puente Formation | North America | ||
| Purisima Formation | North America | ||
| San Mateo Formation | North America | ||
| Santa Margarita Formation | North America | ||
| Temblor Formation | North America | ||
| Topanga Formation | North America | ||
| Bone Valley Formation | North America | ||
| Calvert Formation | North America | ||
| Kirkwood Formation | North America | ||
| North America | |||
| Cojímar Formation | North America | ||
| Kendance Formation | North America | ||
| North America | |||
| Aymamón Limestone | North America | ||
| Almejas Formation | North America | ||
| Carrillo Puerto Formation | North America | ||
| Chagres Formation | North America | ||
| Chucunaque Formation | North America | ||
| Gatún Formation | North America | ||
| Paraná Formation | South America | ||
| Bahía Inglesa Formation | South America | ||
| Castilletes Formation | South America | ||
| Miramar Formation | South America | ||
| Pisco Formation | South America | ||
| Camacho Formation | South America | ||
| Cantaure Formation | South America | ||
| Caujarao Formation | South America | ||
| Socorro Formation | South America | ||
| Urumaco Formation | South America | ||
| Batesford Limestone | Oceania | ||
| Black Rock Sandstone | Oceania | ||
| Gippsland Limestone | Oceania | ||
| Mannum Formation | Oceania | ||
| Morgan Limestone | Oceania | ||
| Port Campbell Limestone | Oceania | ||
| Oceania | |||
| Oceania | |||
| Duho Formation[73] | Asia | ||
| Seogwipo Formation[74] | Asia |
Locations of megalodon fossil discoveries, yellow from the Pliocene and blue from the Miocene[26][71]
Prey relationships

Vertebra of a whale bitten in half by a megalodon with visible gashes from teeth.
Though sharks are generally opportunistic feeders, megalodon’s great size, high-speed swimming capability, and powerful jaws, coupled with an impressive feeding apparatus, made it an apex predator capable of consuming a broad spectrum of animals. Otodus megalodon was probably one of the most powerful predators to have existed.[15] A study focusing on calcium isotopes of extinct and extant elasmobranch sharks and rays revealed that megalodon fed at a higher trophic level than the contemporaneous great white shark («higher up» in the food chain.)[75]
Fossil evidence indicates that megalodon preyed upon many cetacean species, such as dolphins, small whales, cetotheres, squalodontids (shark toothed dolphins), sperm whales, bowhead whales, and rorquals.[51][76][77] In addition to this, they also targeted seals, sirenians, and sea turtles.[70] The shark was an opportunist and piscivorous, and it would have also gone after smaller fish and other sharks.[51] Many whale bones have been found with deep gashes most likely made by their teeth.[30]: 75 Various excavations have revealed megalodon teeth lying close to the chewed remains of whales,[30]: 75 [32] and sometimes in direct association with them.[78]
The feeding ecology of megalodon appears to have varied with age and between sites, like the modern great white shark. It is plausible that the adult megalodon population off the coast of Peru targeted primarily cetothere whales 2.5 to 7 meters (8.2 to 23 ft) in length and other prey smaller than itself, rather than large whales in the same size class as themselves.[76] Meanwhile, juveniles likely had a diet that consisted more of fish.[36][79]
Competition

Megalodon faced a highly competitive environment.[80] Its position at the top of the food chain[81] probably had a significant impact on the structuring of marine communities.[80][82] Fossil evidence indicates a correlation between megalodon and the emergence and diversification of cetaceans and other marine mammals.[30]: 78 [80] Juvenile megalodon preferred habitats where small cetaceans were abundant, and adult megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.[30]: 74–75
Megalodon were contemporaneous with whale-eating toothed whales (particularly macroraptorial sperm whales and squalodontidae), which were also probably among the era’s apex predators, and provided competition.[80] Some attained gigantic sizes, such as Livyatan, estimated between 13.5 to 17.5 meters (44 to 57 ft). Fossilized teeth of an undetermined species of such physeteroids from Lee Creek Mine, North Carolina, indicate it had a maximum body length of 8–10 m and a maximum lifespan of about 25 years. This is very different from similarly sized modern killer whales that live to 65 years, suggesting that unlike the latter, which are apex predators, these physeteroids were subject to predation from larger species such as megalodon or Livyatan.[83] By the Late Miocene, around 11 Mya, macroraptorials experienced a significant decline in abundance and diversity. Other species may have filled this niche in the Pliocene,[80][84] such as the fossil killer whale Orcinus citoniensis which may have been a pack predator and targeted prey larger than itself,[32][85][86][87] but this inference is disputed,[28] and it was probably a generalist predator rather than a marine mammal specialist.[88]
Megalodon may have subjected contemporaneous white sharks to competitive exclusion, as the fossil records indicate that other shark species avoided regions it inhabited by mainly keeping to the colder waters of the time.[89][30]: 77 In areas where their ranges seemed to have overlapped, such as in Pliocene Baja California, it is possible that megalodon and the great white shark occupied the area at different times of the year while following different migratory prey.[30]: 77 [90] Megalodon probably also had a tendency for cannibalism, much like contemporary sharks.[91]
Feeding strategies

Sharks often employ complex hunting strategies to engage large prey animals. Great white shark hunting strategies may be similar to how megalodon hunted its large prey.[92] Megalodon bite marks on whale fossils suggest that it employed different hunting strategies against large prey than the great white shark.[51]
One particular specimen–the remains of a 9-meter (30 ft) long undescribed Miocene baleen whale–provided the first opportunity to quantitatively analyze its attack behavior. Unlike great whites which target the underbelly of their prey, megalodon probably targeted the heart and lungs, with their thick teeth adapted for biting through tough bone, as indicated by bite marks inflicted to the rib cage and other tough bony areas on whale remains.[51] Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans, for example cetotheres, suggest that they were rammed with great force from below before being killed and eaten, based on compression fractures.[92]
There is also evidence that a possible separate hunting strategy existed for attacking raptorial sperm whales; a tooth belonging to an undetermined 4 m (13 ft) physeteroid closely resembling those of Acrophyseter discovered in the Nutrien Aurora Phosphate Mine in North Carolina suggests that a megalodon or O. chubutensis may have aimed for the head of the sperm whale in order to inflict a fatal bite, the resulting attack leaving distinctive bite marks on the tooth. While scavenging behavior cannot be ruled out as a possibility, the placement of the bite marks is more consistent with predatory attacks than feeding by scavenging, as the jaw is not a particularly nutritious area to for a shark feed or focus on. The fact that the bite marks were found on the tooth’s roots further suggest that the shark broke the whale’s jaw during the bite, suggesting the bite was extremely powerful. The fossil is also notable as it stands as the first known instance of an antagonistic interaction between a sperm whale and an otodontid shark recorded in the fossil record.[93]
During the Pliocene, larger cetaceans appeared.[94] Megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones and tail vertebrae of large whales from the Pliocene have been found with megalodon bite marks, which suggests that megalodon would immobilize a large whale before killing and feeding on it.[14][51]
Growth and reproduction

In 2010, Ehret estimated that megalodon had a fast growth rate nearly two times that of the extant great white shark. He also estimated that the slowing or cessation of somatic growth in megalodon occurred around 25 years of age, suggesting that this species had an extremely delayed sexual maturity.[95] In 2021, Shimada and colleagues calculated the growth rate of an approximately 9.2 m (30 ft) individual based on the Belgian vertebrate column specimen that presumably contains annual growth rings on three of its vertebrae. They estimated the individual died at 46 years of age, with a growth rate of 16 cm (6.3 in) per year, and a length of 2 m (6 ft 7 in) at birth. For a 15 m (49 ft) individual—which they considered to have been the maximum size attainable—this would equate to a lifespan of 88 to 100 years.[96] However, Cooper and his colleagues in 2022 estimated the length of this 46 year old individual at nearly 16 m (52 ft) based on the 3D reconstruction which resulted in the complete vertebral column to be 11.1 m (36 ft) long; the researchers claimed that this size estimate difference occurred due to the fact that Shimada and his colleagues extrapolated its size only based on the vertebral centra.[15]
Megalodon, like contemporaneous sharks, made use of nursery areas to birth their young in, specifically warm-water coastal environments with large amounts of food and protection from predators.[36] Nursery sites were identified in the Gatún Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands,[97] and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of megalodon also.[98] Infant megalodons were around 3.5 meters (11 ft) at their smallest,[40]: 61 and the pups were vulnerable to predation by other shark species, such as the great hammerhead shark (Sphyrna mokarran) and the snaggletooth shark (Hemipristis serra).[36] Their dietary preferences display an ontogenetic shift:[40]: 65 Young megalodon commonly preyed on fish,[36] sea turtles,[70] dugongs,[31]: 129 and small cetaceans; mature megalodon moved to off-shore areas and consumed large cetaceans.[30]: 74–75
An exceptional case in the fossil record suggests that juvenile megalodon may have occasionally attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-to-7-meter (13 to 23 ft) long Pliocene shark were found on a rib from an ancestral blue or humpback whale that showed evidence of subsequent healing, which is suspected to have been inflicted by a juvenile megalodon.[99][100]
Extinction
Climate change
The Earth experienced a number of changes during the time period megalodon existed which affected marine life. A cooling trend starting in the Oligocene 35 Mya ultimately led to glaciation at the poles. Geological events changed currents and precipitation; among these were the closure of the Central American Seaway and changes in the Tethys Ocean, contributing to the cooling of the oceans. The stalling of the Gulf Stream prevented nutrient-rich water from reaching major marine ecosystems, which may have negatively affected its food sources. The largest fluctuation of sea levels in the Cenozoic era occurred in the Plio-Pleistocene, between around 5 million to 12 thousand years ago, due to the expansion of glaciers at the poles, which negatively impacted coastal environments, and may have contributed to its extinction along with those of several other marine megafaunal species.[101] These oceanographic changes, in particular the sea level drops, may have restricted many of the suitable shallow warm-water nursery sites for megalodon, hindering reproduction.[102] Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.[103][36]
As its range did not apparently extend into colder waters, megalodon may not have been able to retain a significant amount of metabolic heat, so its range was restricted to shrinking warmer waters.[102][77][104] Fossil evidence confirms the absence of megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[30]: 77 However, an analysis of the distribution of megalodon over time suggests that temperature change did not play a direct role in its extinction. Its distribution during the Miocene and Pliocene did not correlate with warming and cooling trends; while abundance and distribution declined during the Pliocene, megalodon did show a capacity to inhabit colder latitudes. It was found in locations with a mean temperature ranging from 12 to 27 °C (54 to 81 °F), with a total range of 1 to 33 °C (34 to 91 °F), indicating that the global extent of suitable habitat should not have been greatly affected by the temperature changes that occurred.[26] This is consistent with evidence that it was a mesotherm.[49]
Changing ecosystem

Marine mammals attained their greatest diversity during the Miocene,[30]: 71 such as with baleen whales with over 20 recognized Miocene genera in comparison to only six extant genera.[106] Such diversity presented an ideal setting to support a super-predator such as megalodon.[30]: 75 By the end of the Miocene, many species of mysticetes had gone extinct;[80] surviving species may have been faster swimmers and thus more elusive prey.[31]: 46 Furthermore, after the closure of the Central American Seaway, tropical whales decreased in diversity and abundance.[104] The extinction of megalodon correlates with the decline of many small mysticete lineages, and it is possible that it was quite dependent on them as a food source.[76] Additionally, a marine megafauna extinction during the Pliocene was discovered to have eliminated 36% of all large marine species including 55% of marine mammals, 35% of seabirds, 9% of sharks, and 43% of sea turtles. The extinction was selective for endotherms and mesotherms relative to poikilotherms, implying causation by a decreased food supply[101] and thus consistent with megalodon being mesothermic.[49] Megalodon may have been too large to sustain itself on the declining marine food resources.[102] The cooling of the oceans during the Pliocene might have restricted the access of megalodon to the polar regions, depriving it of the large whales which had migrated there.[104]
Competition from large odontocetes, such as macropredatory sperm whales which appeared in the Miocene, and a member of genus Orcinus (i.e., Orcinus citoniensis) in the Pliocene,[80][84] is assumed to have contributed to the decline and extinction of megalodon.[26][31]: 46–47 [102][107] But this assumption is disputed:[28] The Orcininae emerged in Mid-Pliocene with O. citoniensis reported from the
Pliocene of Italy,[84][108] and similar forms reported from the Pliocene of England and South Africa,[84] indicating the capacity of these dolphins to cope with increasingly prevalent cold water temperatures in high latitudes.[84] These dolphins were assumed to have been macrophagous in some studies,[26] but on closer inspection, these dolphins are not found to be macrophagous and fed on small fishes instead.[108] On the other hand, gigantic macropredatory sperm whales such as Livyatan-like forms are last reported from Australia and South Africa circa 5 million years ago.[109][110][111] Others, such as Hoplocetus and Scaldicetus also occupied a niche similar to that of modern killer whales but the last of these forms disappeared during the Pliocene.[112][108] Members of genus Orcinus became large and macrophagous in the Pleistocene.[108]
Paleontologist Robert Boessenecker and his colleagues rechecked the fossil record of megalodon for carbon dating errors and concluded that it disappeared circa 3.5 million years ago.[28] Boessenecker and his colleagues further suggest that megalodon suffered range fragmentation due to climatic shifts,[28] and competition with white sharks might have contributed to its decline and extinction.[28] Competition with white sharks is assumed to be a factor in other studies as well,[113][26][107] but this hypothesis warrants further testing.[114] Multiple compounding environmental and ecological factors including climate change and thermal limitations, collapse of prey populations and resource competition with white sharks are believed to have contributed to decline and extinction of megalodon for now.[107]
The extinction of megalodon set the stage for further changes in marine communities. The average body size of baleen whales increased significantly after its disappearance, although possibly due to other, climate-related, causes.[115] Conversely the increase in baleen whale size may have contributed to the extinction of megalodon, as they may have preferred to go after smaller whales; bite marks on large whale species may have come from scavenging sharks. Megalodon may have simply become coextinct with smaller whale species, such as Piscobalaena nana.[105] The extinction of megalodon had a positive impact on other apex predators of the time, such as the great white shark, in some cases spreading to regions where megalodon became absent.[26][113][116]
In popular culture

HMS Challenger discovered megalodon teeth which were erroneously dated to be around 11,000 to 24,000 years old.
Megalodon has been portrayed in many works of fiction, including films and novels, and continues to be a popular subject for fiction involving sea monsters.[117] Reports of supposedly fresh megalodon teeth, such as those found by HMS Challenger in 1873 which were dated in 1959 by the zoologist Wladimir Tschernezky to be around 11,000 to 24,000 years old, helped popularise claims of recent megalodon survival amongst cryptozoologists.[118] These claims are now discredited, and are probably teeth that were well-preserved by a thick mineral-crust precipitate of manganese dioxide, and so had a lower decomposition rate and retained a white color during fossilization. Fossil megalodon teeth can vary in color from off-white to dark browns and greys, and some fossil teeth may have been redeposited into a younger stratum. The claims that megalodon could remain elusive in the depths, similar to the megamouth shark which was discovered in 1976, are unlikely as the shark lived in warm coastal waters and probably could not survive in the cold and nutrient-poor deep sea environment.[119][120]
Contemporary fiction about megalodon surviving into modern times was pioneered by the 1997 novel Meg: A Novel of Deep Terror by Steve Alten and its subsequent sequels. Megalodon subsequently began to feature in films, such as the 2003 direct to video Shark Attack 3: Megalodon, and later The Meg, a 2018 film based on the 1997 book which grossed over $500 million at the box office.[118][121]
Animal Planet’s pseudo-documentary Mermaids: The Body Found included an encounter 1.6 mya between a pod of mermaids and a megalodon.[122] Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives,[123] a controversial docufiction about the creature that presented alleged evidence in order to suggest that megalodons still lived. This program received criticism for being completely fictional and for inadequately disclosing its fictional nature; for example, all of the supposed scientists depicted were paid actors, and there was no disclosure in the documentary itself that it was fictional. In a poll by Discovery, 73% of the viewers of the documentary thought that megalodon was not extinct. In 2014, Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.[51][124][125][126] Despite the criticism from scientists, Megalodon: The Monster Shark Lives was a huge ratings success, gaining 4.8 million viewers, the most for any Shark Week episode up to that point.[127]
Megalodon teeth are the state fossil of North Carolina.[128]
See also
- List of prehistoric cartilaginous fish
- Prehistoric fish
- Largest prehistoric organisms
Notes
- ^ Carbonated bioapatite from a megalodon tooth (of unknown source location) dated to 5.75 ± 0.9 Ma in age has been analyzed for isotope ratios of oxygen (18O/16O) and carbon (13C/12C), using a carbonate clumped-isotope thermometer methodology to yield an estimate of the ambient temperature in that individual’s environment of 19 ± 4 °C.[69]
References
- ^ a b Agassiz, Louis (1843). Recherches sur les poissons fossiles [Research on the fossil fishes] (in French). Neuchatel: Petitpierre. p. 41.
- ^ «Otodus (Megaselachus) megalodon (Agassiz, 1837)». SharkReferences.com. Retrieved 24 October 2017.
- ^ Eastman, C. R. (1904). Maryland Geological Survey. Vol. 2. Baltimore, Maryland: Johns Hopkins University. p. 82.
- ^ a b c Cappetta, H. (1987). «Mesozoic and Cenozoic Elasmobranchii». Handbook of Paleoichthyology. Vol. 3B. München, Germany: Friedrich Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- ^ Hay, O. P. (1901). «Bibliography and Catalogue of the Fossil Vertebrata of North America». Bulletin of the United States Geological Society (179): 308.
- ^ a b c d e f Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (2016). «A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade». Historical Biology. 29 (5): 1–11. doi:10.1080/08912963.2016.1236795. S2CID 89080495.
- ^ a b c d e f Shimada, Kenshu (2019). «The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited». Historical Biology. 33 (7): 1–8. doi:10.1080/08912963.2019.1666840. ISSN 0891-2963. S2CID 208570844.
- ^ a b Cooper, J. A.; Pimiento, C.; Ferrón, H. G.; Benton, M. J. (2020). «Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction». Scientific Reports. 10 (14596): 14596. Bibcode:2020NatSR..1014596C. doi:10.1038/s41598-020-71387-y. PMC 7471939. PMID 32883981.
- ^ «Giant ‘megalodon’ shark extinct earlier than previously thought». Science Daily. 13 February 2019.
- ^ a b c d e Perez, Victor; Leder, Ronny; Badaut, Teddy (2021). «Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions». Palaeontologia Electronica. 24 (1): 1–28. doi:10.26879/1140.
- ^ a b Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c d e Pimiento, C.; Balk, M. A. (2015). «Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators». Paleobiology. 41 (3): 479–490. doi:10.1017/pab.2015.16. PMC 4541548. PMID 26321775.
- ^ a b c Prothero, Donald R. (25 August 2015). «09. Mega-Jaws: The Largest Fish. Carcharocles». The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution. New York Chichester, West Sussex: Columbia University Press. pp. 96–110. doi:10.7312/prot17190-010. ISBN 978-0-231-53942-5.
- ^ a b c d e Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?» (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ^ a b c d e f Cooper, J.A.; Hutchinson, J.R.; Bernvi, D.C.; Cliff, G.; Wilson, R.P.; Dicken, M.L.; Menzel, J.; Wroe, S.; Pirlo, J.; Pimiento, C. (2022). «The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling». Science Advances. 8 (33): eabm9424. Bibcode:2022SciA….8M9424C. doi:10.1126/sciadv.abm9424. ISSN 2375-2548. PMC 9385135. PMID 35977007.
- ^ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Westport, Connecticut: Libraries Unlimited. pp. 25–26. ISBN 978-1-59158-265-6. OCLC 230807846.
- ^ Hsu, Kuang-Tai (2009). «The Path to Steno’s Synthesis on the Animal Origin of Glossopetrae». In Rosenburg, G. D. (ed.). The Revolution in Geology from the Renaissance to the Enlightenment. Vol. 203. Boulder, Colorado: Geological Society of America. ISBN 978-0-8137-1203-1. OCLC 608657795.
- ^ Eilperin, J. (2012). Demon Fish. Pantheon Books. p. 43. ISBN 978-0-7156-4352-5.
- ^ a b c d e f Nyberg, K. G.; Ciampaglio C. N.; Wray G. A. (2006). «Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth». Journal of Vertebrate Paleontology. 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. S2CID 53640614.
- ^ Keyes, I. W. (2012). «New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record». New Zealand Journal of Geology and Geophysics. 15 (2): 229. doi:10.1080/00288306.1972.10421956.
- ^ μέγας. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ ὀδούς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ Lawley, R. (1881). «Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)» [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (in Italian). 5: 167–172.
- ^ Yabe, H.; Goto, M.; Kaneko, N. (2004). «Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records». The Palaeontological Society of Japan. 75: 7–15.
- ^ a b c Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ a b c d e f g h i j k l Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c Pimiento, C.; Clements, C. F. (2014). «When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record». PLOS ONE. 9 (10): e111086. Bibcode:2014PLoSO…9k1086P. doi:10.1371/journal.pone.0111086. PMC 4206505. PMID 25338197.
- ^ a b c d e f Boessenecker, R. W.; Ehret, D. J.; Long, D. J.; Churchill, M.; Martin, E.; Boessenecker, S. J. (2019). «The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific». PeerJ. 7: e6088. doi:10.7717/peerj.6088. PMC 6377595. PMID 30783558.
- ^ a b c d Ehret D. J.; Hubbell G.; Macfadden B. J. (2009). «Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru». Journal of Vertebrate Paleontology. 29 (1): 1–13. doi:10.1671/039.029.0113. JSTOR 20491064. S2CID 129585445.
- ^ a b c d e f g h i j k l m n o p q r Klimley, Peter; Ainley, David (1996). «Evolution». Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. ISBN 978-0-12-415031-7. OCLC 212425118.
- ^ a b c d e f g h i j k l m n o p Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ a b c d e Andres, Lutz (2002). «C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles«. fossilguy.com. Retrieved 16 January 2008.
- ^ a b Perez, V. J.; Godfrey, S. J.; Kent, B. W.; Weems, R. E.; Nance, J. R. (2019). «The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time». Journal of Vertebrate Paleontology. 38 (6): e1546732. doi:10.1080/02724634.2018.1546732.
- ^ Siverson, M.; Lindgren, J.; Newbrey, M.G.; Cederström, P.; Cook, T.D. (2013). «Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type» (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. S2CID 58906204. Archived from the original (PDF) on 19 October 2013.
- ^ Benton, M. J.; Pearson, P. N. (2001). «Speciation in the fossil record». Trends in Ecology and Evolution. 16 (7): 405–411. doi:10.1016/s0169-5347(01)02149-8. PMID 11403874.
- ^ a b c d e f g h Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Vivian, G. (2013). «Research debunks Great White lineage».
- ^ «Palaeontological detective work unravels evolution of megatooth sharks». 2013.
- ^ a b c d e f g h i j k l m n o p q r Gottfried, MD; Compagno, LJV; Bowman, SC (1996). «Size and skeletal anatomy of the giant megatooth shark Carcharodon megalodon«. In Klimley; Ainley (eds.). Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. pp. 55–89. ISBN 978-0124150317.
- ^ «Could Megalodon Have Looked Like a BIG Sandtiger Shark?». Biology of Sharks and Rays. Retrieved 2 September 2017.
- ^ Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). «Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles» (PDF). Caribbean Journal of Science. 44 (3): 279–286. doi:10.18475/cjos.v44i3.a2. S2CID 87154947. Archived from the original (PDF) on 20 July 2011.
- ^ Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ a b McClain, Craig R.; Balk, Meghan A.; Benfield, Mark C.; Branch, Trevor A.; Chen, Catherine; Cosgrove, James; Dove, Alistair D.M.; Gaskins, Lindsay C.; Helm, Rebecca R.; Hochberg, Frederick G.; Lee, Frank B. (13 January 2015). «Sizing ocean giants: patterns of intraspecific size variation in marine megafauna». PeerJ. 3: e715. doi:10.7717/peerj.715. ISSN 2167-8359. PMC 4304853. PMID 25649000.
- ^ I., Castro, Jose (2011). Sharks of North America. Oxford University Press, USA. ISBN 978-0-19-978097-6. OCLC 958576172.
- ^ Borrell, Asunción; Aguilar, Alex; Gazo, Manel; Kumarran, R. P.; Cardona, Luis (1 December 2011). «Stable isotope profiles in whale shark (Rhincodon typus) suggest segregation and dissimilarities in the diet depending on sex and size». Environmental Biology of Fishes. 92 (4): 559–567. doi:10.1007/s10641-011-9879-y. ISSN 1573-5133. S2CID 37683420.
- ^ Jacoby, D. M. P.; Siriwat, P.; Freeman, R.; Carbone, C. (2015). «Is the scaling of swim speed in sharks driven by metabolism?». Biology Letters. 12 (10): 20150781. doi:10.1098/rsbl.2015.0781. PMC 4707698. PMID 26631246.
- ^ a b c Ferrón, H. G. (2017). «Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks». PLOS ONE. 12 (9): e0185185. Bibcode:2017PLoSO..1285185F. doi:10.1371/journal.pone.0185185. PMC 5609766. PMID 28938002.
- ^ Shimada, K.; Becker, M. A.; Griffiths, M. L. (2020). «Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to ‘off-the-scale’ gigantism of the megatooth shark, Otodus megalodon«. Historical Biology. 33 (11): 1–17. doi:10.1080/08912963.2020.1812598.
- ^ a b c d e f g Prothero, D. R. (2015). «Mega-Jaws». The Story of Life in 25 Fossils. New York, New York: Columbia University Press. pp. 96–110. ISBN 978-0-231-17190-8. OCLC 897505111.
- ^ Helfman, G.; Burgess, G. H. (2014). Sharks: The Animal Answer Guide. Baltimore, Maryland: Johns Hopkins University Press. p. 19. ISBN 978-1-4214-1310-5. OCLC 903293986.
- ^ Randall, John E. (1973). «Size of the Great White Shark (Carcharodon)». Science Magazine. 181 (4095): 169–170. Bibcode:1973Sci…181..169R. doi:10.1126/science.181.4095.169. PMID 17746627. S2CID 36607712.
- ^ Schembri, Patrick (1994). «Malta’s Natural Heritage». Natural Heritage. In: 105–124.
- ^ Papson, Stephen (1992). «Copyright: Cross the Fin Line of Terror». Journal of American Culture. 15 (4): 67–81. doi:10.1111/j.1542-734X.1992.1504_67.x.
- ^ Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ Shimada, Kenshu (2002). «The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)». Journal of Fossil Research. 35 (2): 28–33.
- ^ a b Pimiento, Catalina; Gerardo González-Barba; Dana J. Ehret; Austin J. W. Hendy; Bruce J. MacFadden; Carlos Jaramillo (2013). «Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama» (PDF). Journal of Paleontology. 87 (5): 755–774. doi:10.1666/12-117. S2CID 45662900. Archived from the original (PDF) on 29 October 2013.
- ^ Crane, B. (2017). «A Prehistoric Killer, Buried in Muck». The New Yorker. Retrieved 10 December 2017.
- ^ Mustain, A. (2011). «For Sale: World’s Largest Shark Jaws». LiveScience. Retrieved 31 August 2017.
- ^ a b Almgreen, S. E. Bendix (15 November 1983). «Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn». Bulletin of the Geological Society of Denmark. 32: 1–32. CiteSeerX 10.1.1.514.1782. doi:10.37570/bgsd-1983-32-01. S2CID 53311833. NAID 10012345550.
- ^ Reolid, M.; Molina, J. M. (2015). «Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain)». Estudios Geológicos. 71 (2): e032. doi:10.3989/egeol.41828.342.
- ^ Uyeno, T.; Sakamoto, O.; Sekine, H. (1989). «The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan». Saitama Museum of Natural History Bulletin. 7: 73–85.
- ^ Anderson, P.S.L.; Westneat, M. (2009). «A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)». Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1. S2CID 86203770.
- ^ Ballell, A.; Ferrón, H.G. (2021). «Biomechanical insights into the dentition of megatooth sharks (Lamniformes: Otodontidae)». Scientific Reports. 11 (1232): 1232. doi:10.1038/s41598-020-80323-z. PMC 7806677. PMID 33441828.
- ^ a b Stringer, G. L.; King, L. (2012). «Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana». New Mexico Museum of Natural History and Science, Bulletin. Vertebrate Corpolites. 57: 301.
- ^ «Megalodon Shark Facts and Information: The Details». fossilguy.com. Retrieved 18 September 2017.
- ^ Fitzgerald, Erich (2004). «A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia». Memoirs of Museum Victoria. 61 (2): 183–208. doi:10.24199/j.mmv.2004.61.12.
- ^ Löffler, N.; Fiebig, J.; Mulch, A.; Tütken, T.; Schmidt, B.C.; Bajnai, D.; Conrad, A.C.; Wacker, U.; Böttcher, M.E. (2019). «Refining the temperature dependence of the oxygen and clumped isotopic compositions of structurally bound carbonate in apatite». Geochimica et Cosmochimica Acta. 253: 19–38. Bibcode:2019GeCoA.253…19L. doi:10.1016/j.gca.2019.03.002. S2CID 107992832.
- ^ a b c Aguilera O.; Augilera E. R. D. (2004). «Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage». Caribbean Journal of Science. 40 (3): 362–368.
- ^ a b «Carcharocles megalodon«. Fossilworks. Retrieved 28 August 2017 from the Paleobiology Database.
{{cite web}}: CS1 maint: postscript (link) - ^ «Young Prince George gifted 23m-year-old tooth from extinct shark found in Malta». Times of Malta. 27 September 2020. Retrieved 28 September 2020.
- ^ Yun, C. (2020). «New example of Cosmopolitodus hastalis (Lamniformes, Lamnidae) from the Miocene South Korea». Zoodiversity. 54 (5): 433–438. doi:10.15407/zoo2020.05.433. S2CID 229274996.
- ^ Choi, Seung; Lee, Yuong-Nam (2017). «A review of vertebrate body fossils from the Korean Peninsula and perspectives». Geosciences Journal. 21 (6): 867–889. Bibcode:2017GescJ..21..867C. doi:10.1007/s12303-017-0040-6. ISSN 1226-4806. S2CID 133835817.
- ^ Martin, J. E.; Tacail, T.; Sylvain, A.; Catherine, G.; Vincent, B. (2015). «Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs». Chemical Geology. 415: 118–125. Bibcode:2015ChGeo.415..118M. doi:10.1016/j.chemgeo.2015.09.011.
- ^ a b c Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ a b Morgan, Gary S. (1994). «Whither the giant white shark?» (PDF). Paleontology Topics. 2 (3): 1–2. Archived from the original (PDF) on 22 July 2016.
- ^ Augilera, Orangel A.; García, Luis; Cozzuol, Mario A. (2008). «Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation». Paläontologische Zeitschrift. 82 (2): 204–208. doi:10.1007/BF02988410. ISSN 0038-2353. S2CID 84251638.
- ^ Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). «The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)». Journal of South American Earth Sciences. 73: 168–190. Bibcode:2017JSAES..73..168L. doi:10.1016/j.jsames.2016.12.010.
- ^ a b c d e f g h Lambert, O.; Bianucci, G.; Post, P.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. (2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020. S2CID 4369352.
- ^ Compagno, Leonard J. V. (1989). «Alternative life-history styles of cartilaginous fishes in time and space». Environmental Biology of Fishes. 28 (1–4): 33–75. doi:10.1007/BF00751027. S2CID 22527888.
- ^ Ferretti, Francesco; Boris Worm; Gregory L. Britten; Michael R. Heithaus; Heike K. Lotze1 (2010). «Patterns and ecosystem consequences of shark declines in the ocean» (PDF). Ecology Letters. 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. Archived from the original (PDF) on 6 July 2011. Retrieved 19 February 2011.
- ^ Gilbert, K.N.; Ivany, L.C.; Uhen, M.D. (2018). «Living fast and dying young: life history and ecology of a Neogene sperm whale». Journal of Vertebrate Paleontology. 38 (2): e1439038. doi:10.1080/02724634.2018.1439038. S2CID 89750852.
- ^ a b c d e Heyning, John; Dahlheim, Marilyn (1988). «Orcinus orca» (PDF). Mammalian Species (304): 1–9. doi:10.2307/3504225. JSTOR 3504225. S2CID 253914153. Archived from the original (PDF) on 5 December 2010.
- ^ Bianucci, Giovanni; Walter, Landini (2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society. 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x.
- ^ Lindberg, D. R.; Pyenson, N. D. (2006). «Evolutionary Patterns in Cetacea: Fishing Up Prey Size through Deep Time». Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 77. ISBN 978-0-520-24884-7.
- ^ Boessenecker, R. W. (2013). «A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans». Geodiversitas. 35 (4): 815–940. doi:10.5252/g2013n4a5. S2CID 85940452.
- ^ Bianucci, G. (1997). «Hemisyntrachelus cortesii (Cetacea, Delphinidae) from the Pliocene Sediments of Campore Quarry (Salsomaggiori Terme, Italy». Bollettino della Societa Paleontologica Italiana. 36 (1): 75–83).
- ^ Antunes, M.T.; Legoinha, P.; Balbing, A. (2015). «Megalodon, mako shark and planktonic foraminifera from the continental shelf off Portugal and their age». Geologica Acta. 13: 181–190.
- ^ «Paleoecology of Megalodon and the White Shark». Biology of Sharks and Rays. Retrieved 1 October 2017.
- ^ Tanke, Darren; Currie, Philip (1998). «Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence» (PDF). Gaia (15): 167–184.
- ^ a b Godfrey, S. J.; Altman, J. (2005). «A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon» (PDF). Jeffersoniana (16): 1–12.
- ^ STEPHEN J. GODFREY; JOHN R. NANCE; NORMAN L. RIKER (2021). «Otodus-bitten sperm whale tooth from the Neogene of the Coastal Eastern United States» (PDF). Acta Palaeontologica Polonica. 66 (3): 599–603.
- ^ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). «The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes». Journal of Mammalian Evolution. 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3. S2CID 90231.
- ^ Ehret D.J. (2010). «CHAPTER 5 – MACROEVOLUTION, AGE, AND GROWTH DETERMINATION OF THE MEGATOOTHED SHARKS (LAMNIFORMES: OTODONTIDAE)». Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes) (PDF). pp. 100–136.
- ^ S., Kenshu; Bonnan, M. F.; Becker, M. A.; Griffiths, M. L. (2021). «Ontogenetic growth pattern of the extinct megatooth shark Otodus megalodon—implications for its reproductive biology, development, and life expectancy». Historical Biology. 33 (12): 3254–3259. doi:10.1080/08912963.2020.1861608.
- ^ «Identifican en Canarias fósiles de ‘megalodón’, el tiburón más grande que ha existido» [Identifying Canary fossils of ‘megalodon’, the largest shark that ever lived] (in Spanish). Europa Press Noticias SA. 2013. Retrieved 29 August 2017.
- ^ Dulvy, N. K.; Reynolds, J. D. (1997). «Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays». Proceedings of the Royal Society B: Biological Sciences. 264 (1386): 1309–1315. Bibcode:1997RSPSB.264.1309D. doi:10.1098/rspb.1997.0181. PMC 1688595.
- ^ Godfrey, Stephen (2004). «The Ecphora» (PDF). The Newsletter of Calvert Marine Museum Fossil Club. 19 (1): 1–13. Archived from the original (PDF) on 10 December 2010.
- ^ Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (27 August 2010). «Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event». International Journal of Osteoarchaeology. 22 (3): 253–260. doi:10.1002/oa.1199.
- ^ a b Pimiento, C.; Griffin, J. N.; Clements, C. F.; Silvestro, D.; Varela, S.; Uhen, M. D.; Jaramillo, C. (2017). «The Pleistocene Marine Megafauna Extinction and its Impact on Functional Diversity». Nature Ecology and Evolution. 1 (8): 1100–1106. doi:10.1038/s41559-017-0223-6. PMID 29046566. S2CID 3639394.
- ^ a b c d «The Extinction of Megalodon». Biology of Sharks and Rays. Retrieved 31 August 2017.
- ^ Reilly, Michael (29 September 2009). «Prehistoric Shark Nursery Spawned Giants». Discovery News. Archived from the original on 10 March 2012. Retrieved 23 November 2013.
- ^ a b c Allmon, Warren D.; Steven D. Emslie; Douglas S. Jones; Gary S. Morgan (2006). «Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms». The Journal of Geology. 104 (2): 143–162. Bibcode:1996JG….104..143A. doi:10.1086/629811. S2CID 128418299.
- ^ a b Collareta, A.; Lambert, O.; Landini, W.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ Dooly A.C.; Nicholas C. F.; Luo Z. X. (2006). «The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)». Journal of Vertebrate Paleontology. 24 (2): 453–463. doi:10.1671/2401. JSTOR 4524731. S2CID 84970052.
- ^ a b c McCormack, Jeremy; Griffiths, Michael L.; Kim, Sora L.; Shimada, Kenshu; Karnes, Molly; Maisch, Harry; Pederzani, Sarah; Bourgon, Nicolas; Jaouen, Klervia; Becker, Martin A.; Jöns, Niels (31 May 2022). «Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes». Nature Communications. 13 (1): 2980. Bibcode:2022NatCo..13.2980M. doi:10.1038/s41467-022-30528-9. ISSN 2041-1723. PMC 9156768. PMID 35641494. S2CID 249235478.
- ^ a b c d Citron, Sara; Geisler, Jonathan H.; Alberto, Collareta; Giovanni, Bianucci (2022). «Systematics, phylogeny and feeding behavior of the oldest killer whale: a reappraisal of Orcinus citoniensis (Capellini, 1883) from the Pliocene of Tuscany (Italy)». Bollettino della Società Paleontologica Italiana. 61 (2): 167–186. doi:10.4435/BSPI.2022.13.
- ^ «Huge Tooth Reveals Prehistoric Moby Dick in Melbourne». Australasian Science Magazine. Retrieved 24 April 2016.
- ^ «Move over Moby Dick: Meet Melbourne’s own mega whale». The Sydney Morning Herald. 21 April 2016.
- ^ Govender, R (2021). «Early Pliocene fossil cetaceans from Hondeklip Bay, Namaqualand, South Africa». Historical Biology. 33 (4): 574–593. doi:10.1080/08912963.2019.1650273. S2CID 202019648.
- ^ Hampe, O. (2006). «Middle/late Miocene hoplocetine sperm whale remains (Odontoceti: Physeteridae) of North Germany with an emended classification of the Hoplocetinae». Fossil Record. 9 (1): 61–86. doi:10.1002/mmng.200600002.
- ^ a b Antunes, Miguel Telles; Balbino, Ausenda Cáceres (2010). «The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic». Revista Española de Paleontología. 25 (1): 1–6.
- ^ Kast, Emma R.; Griffiths, Michael L.; Kim, Sora. L.; Rao, Zixuan C.; Shimada, Kensu; Becker, Martin A.; Maisch, Harry M.; Eagle, Robert A.; Clarke, Chelesia A.; Neumann, Allison N.; Karnes, Molly E.; Lüdecke, Tina; Leichliter, Jennifer N.; Martínez-García, Alfredo; Akhtar, Alliya A.; Wang, Xingchen T.; Haug, Gerald H.; Sigman, Daniel M. (22 June 2022). «Cenozoic megatooth sharks occupied extremely high trophic positions». Science Advances. 8 (25): eabl6529. Bibcode:2022SciA….8L6529K. doi:10.1126/sciadv.abl6529. PMC 9217088. PMID 35731884.
- ^ Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (2017). «Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics». Proceedings of the Royal Society B: Biological Sciences. 284 (1855): 20170546. doi:10.1098/rspb.2017.0546. PMC 5454272. PMID 28539520.
- ^ Sylvain, Adnet; A. C. Balbino; M. T. Antunes; J. M. Marín-Ferrer (2010). «New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean». Neues Jahrbuch für Geologie und Paläontologie. 256 (1): 7–16. doi:10.1127/0077-7749/2009/0029.
- ^ Weinstock, J. A. (2014). The Ashgate Encyclopedia of Literary and Cinematic Monsters. Farnham, United Kingdom: Routledge. pp. 107–108. ISBN 978-1-4094-2562-5. OCLC 874390267.
- ^ a b Guimont, Edward (5 October 2021). «The Megalodon: A Monster of the New Mythology». M/C Journal. 24 (5). doi:10.5204/mcj.2793. ISSN 1441-2616. S2CID 241813307.
- ^ Roesch, B. S. (1998). «A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles megalodon«. The Cryptozoology Review. 3 (2): 14–24.
- ^ «Does Megalodon Still Live?». Biology of Sharks and Rays. Retrieved 2 October 2017.
- ^ «The Meg (2018)». Box Office Mojo. Retrieved 22 December 2017.
- ^ Sid Bennett (director) (27 May 2012). Mermaids: The Body Found (Motion picture). Animal Planet.
- ^ «Shark Week ‘Megalodon: The Monster Shark Lives’ Tries To Prove Existence Of Prehistoric Shark (VIDEO)». Huff Post Green. 5 August 2013. Retrieved 11 August 2013.
- ^ Winston, B.; Vanstone, G.; Chi, W. (2017). «A Walk in the Woods». The Act of Documenting: Documentary Film in the 21st Century. New York, New York: Bloomsbury Publishing. ISBN 978-1-5013-0918-2. OCLC 961183719.
- ^ Flanagin, J. (2014). «Sorry, Fans. Discovery Has Jumped the Shark Week». New York Times. Retrieved 16 August 2014.
- ^ Shiffman, David (15 August 2014). «Shark Week Is Lying Again About Monster Megalodon Sharks». Slate Magazine. Retrieved 31 July 2022.
- ^ O’Connell, Mikey (5 August 2013). «TV Ratings: Shark Week Hits Record Highs With Fake ‘Megalodon’ Doc». The Hollywood Reporter. Retrieved 31 July 2022.
- ^ «Fossil, Fossilized Teeth of the Megalodon Shark | NCpedia». ncpedia.org. Retrieved 17 October 2019.
Further reading
- Dickson, K. A.; Graham, J. B. (November–December 2004). «Evolution and consequences of endothermy in fishes». Physiological and Biochemical Zoology. 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772. S2CID 40104003.
- Kent, Bretton W. (1994). Fossil Sharks of the Chesapeake Bay Region. Columbia, Md.: Egan Rees & Boyer. ISBN 978-1-881620-01-3. OCLC 918266672.
External links
- The rise of super predatory sharks
- Carcharocles: Extinct Megatoothed shark
- Paleontological videos
- Paleontologist Mark Renz shows one of the largest megalodon teeth discovered on YouTube
- Shark Week Special on megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about megalodon on YouTube (featuring expert Dana Ehret)
- Lamniform sharks: 110 million years of ocean supremacy on YouTube (featuring expert Mikael Siverson)
- The Rise and Fall of the Neogene Giant Sharks on YouTube (featuring expert Bretton Kent)
- Moore, Kallie (19 December 2018). «Why Megalodon (Definitely) Went Extinct». PBS Eons. Archived from the original on 14 November 2021 – via YouTube.
| Megalodon
Temporal range: Early Miocene–Early Pliocene, c. 23–3.6 Ma PreꞒ Ꞓ O S D C P T J K Pg N |
|
|---|---|
|
|
|
| Model of megalodon jaws at the American Museum of Natural History | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Otodontidae |
| Genus: | †Otodus |
| Species: |
†O. megalodon |
| Binomial name | |
| †Otodus megalodon
(Agassiz, 1843)[1] |
|
| Synonyms[2][3][4][5][6] | |
|
List of synonyms
|
Megalodon (Otodus megalodon),[6][7][8] meaning «big tooth», is an extinct species of mackerel shark that lived approximately 23 to 3.6 million years ago (Mya), from the Early Miocene to the Pliocene epochs.[9] It was formerly thought to be a member of the family Lamnidae and a close relative of the great white shark (Carcharodon carcharias). However, it is now classified into the extinct family Otodontidae, which diverged from the great white shark during the Early Cretaceous.
While regarded as one of the largest and most powerful predators to have ever lived, the megalodon is only known from fragmentary remains, and its appearance and maximum size are uncertain. Scientists differ on whether it would have more closely resembled a stockier version of the great white shark, the whale shark (Rhincodon typus), the basking shark (Cetorhinus maximus) or the sand tiger shark (Carcharias taurus). The most recent estimate with the least error range suggests a maximum length estimate up to 20.3 meters (67 ft),[10] although the modal lengths are estimated at 10.5 meters (34 ft).[11][12] Extrapolation from a vertebral centra with dimensions based on the great white shark suggests that a megalodon about 16 meters (52 ft) long weighs up to 48 metric tons (53 short tons), 17 meters (56 ft) long weighs up to 59 metric tons (65 short tons), and 20.3 meters (67 ft) long (the maximum length) weighs up to 103 metric tons (114 short tons).[13][14] Extrapolating from a vertebral column and reconstructing a 3D model with dimensions based on all extant lamnid sharks suggests that a 16-meter-long (52 ft) individual may have been much larger than previous estimates, reaching an excess of 61.5 metric tons (67.8 short tons) in body mass; an individual of this size would have needed to consume 98,175 kcal per day.[15] Their teeth were thick and robust, built for grabbing prey and breaking bone, and their large jaws could exert a bite force of up to 108,500 to 182,200 newtons (24,400 to 41,000 lbf).[14]
Megalodon probably had a major impact on the structure of marine communities. The fossil record indicates that it had a cosmopolitan distribution. It probably targeted large prey, such as whales, seals and sea turtles. Juveniles inhabited warm coastal waters and fed on fish and small whales. Unlike the great white, which attacks prey from the soft underside, megalodon probably used its strong jaws to break through the chest cavity and puncture the heart and lungs of its prey.
The animal faced competition from whale-eating cetaceans, such as Livyatan and other macroraptorial sperm whales and possibly smaller ancestral killer whales. As the shark preferred warmer waters, it is thought that oceanic cooling associated with the onset of the ice ages, coupled with the lowering of sea levels and resulting loss of suitable nursery areas, may have also contributed to its decline. A reduction in the diversity of baleen whales and a shift in their distribution toward polar regions may have reduced megalodon’s primary food source. The shark’s extinction coincides with a gigantism trend in baleen whales.
Taxonomy
Naming
The depiction of a shark’s head by Nicolas Steno in his work The Head of a Shark Dissected
According to Renaissance accounts, gigantic triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolas Steno, who recognized them as shark teeth, and famously produced a depiction of a shark’s head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a megalodon tooth.[16][17][18]
Swiss naturalist Louis Agassiz gave this shark its initial scientific name, Carcharodon megalodon, in his 1843 work Recherches sur les poissons fossiles, based on tooth remains.[1][19] English paleontologist Edward Charlesworth in his 1837 paper used the name Carcharias megalodon, while citing Agassiz as the author, indicating that Agassiz described the species prior to 1843. English paleontologist Charles Davies Sherborn in 1928 listed an 1835 series of articles by Agassiz as the first scientific description of the shark.[20] The specific name megalodon translates to «big tooth», from Ancient Greek: μέγας, romanized: (mégas), lit. ‘big, mighty’ and ὀδούς (odoús), «tooth».[21][22] The teeth of megalodon are morphologically similar to those of the great white shark (Carcharodon carcharias), and on the basis of this observation, Agassiz assigned megalodon to the genus Carcharodon.[19]
There was one apparent description of the shark in 1881 classifying it as Selache manzonii.[23]
Evolution
While the earliest megalodon remains have been reported from the Late Oligocene, around 28 million years ago (Mya),[24][25] there is disagreement as to when it appeared, with dates ranging to as young as 16 mya.[26] It has been thought that megalodon became extinct around the end of the Pliocene, about 2.6 Mya;[26][27] claims of Pleistocene megalodon teeth, younger than 2.6 million years old, are considered unreliable.[27] A 2019 assessment moves the extinction date back to earlier in the Pliocene, 3.6 Mya.[28]
Megalodon is now considered to be a member of the family Otodontidae, genus Otodus, as opposed to its previous classification into Lamnidae, genus Carcharodon.[26][12][27][6][7] Megalodon’s classification into Carcharodon was due to dental similarity with the great white shark, but most authors currently believe that this is due to convergent evolution. In this model, the great white shark is more closely related to the extinct broad-toothed mako (Isurus hastalis) than to megalodon, as evidenced by more similar dentition in those two sharks; megalodon teeth have much finer serrations than great white shark teeth. The great white shark is more closely related to the mako shark (Isurus spp.), with a common ancestor around 4 Mya.[19][29] Proponents of the former model, wherein megalodon and the great white shark are more closely related, argue that the differences between their dentition are minute and obscure.[30]: 23–25
The genus Carcharocles currently contains four species: C. auriculatus, C. angustidens, C. chubutensis, and C. megalodon.[31]: 30–31 The evolution of this lineage is characterized by the increase of serrations, the widening of the crown, the development of a more triangular shape, and the disappearance of the lateral cusps.[31]: 28–31 [32] The evolution in tooth morphology reflects a shift in predation tactics from a tearing-grasping bite to a cutting bite, likely reflecting a shift in prey choice from fish to cetaceans.[33] Lateral cusplets were finally lost in a gradual process that took roughly 12 million years during the transition between C. chubutensis and C. megalodon.[33] The genus was proposed by D. S. Jordan and H. Hannibal in 1923 to contain C. auriculatus. In the 1980s, megalodon was assigned to Carcharocles.[19][31]: 30 Before this, in 1960, the genus Procarcharodon was erected by French ichthyologist Edgard Casier, which included those four sharks and was considered separate from the great white shark. It is now considered a junior synonym of Carcharocles.[31]: 30 The genus Palaeocarcharodon was erected alongside Procarcharodon to represent the beginning of the lineage, and, in the model wherein megalodon and the great white shark are closely related, their last common ancestor. It is believed to be an evolutionary dead-end and unrelated to the Carcharocles sharks by authors who reject that model.[30]: 70
The great white shark (Carcharodon carcharias) and megalodon were previously thought to be close relatives.[19][29]
Another model of the evolution of this genus, also proposed by Casier in 1960, is that the direct ancestor of the Carcharocles is the shark Otodus obliquus, which lived from the Paleocene through the Miocene epochs, 60 to 13 Mya.[29][32] The genus Otodus is ultimately derived from Cretolamna, a shark from the Cretaceous period.[6][34] In this model, O. obliquus evolved into O. aksuaticus, which evolved into C. auriculatus, and then into C. angustidens, and then into C. chubutensis, and then finally into C. megalodon.
Another model of the evolution of Carcharocles, proposed in 2001 by paleontologist Michael Benton, is that the three other species are actually a single species of shark that gradually changed over time between the Paleocene and the Pliocene, making it a chronospecies.[31]: 17 [25][35] Some authors suggest that C. auriculatus, C. angustidens, and C. chubutensis should be classified as a single species in the genus Otodus, leaving C. megalodon the sole member of Carcharocles.[25][36]
The genus Carcharocles may be invalid, and the shark may actually belong in the genus Otodus, making it Otodus megalodon.[4] A 1974 study on Paleogene sharks by Henri Cappetta erected the subgenus Megaselachus, classifying the shark as Otodus (Megaselachus) megalodon, along with O. (M.) chubutensis. A 2006 review of Chondrichthyes elevated Megaselachus to genus, and classified the sharks as Megaselachus megalodon and M. chubutensis.[4] The discovery of fossils assigned to the genus Megalolamna in 2016 led to a re-evaluation of Otodus, which concluded that it is paraphyletic, that is, it consists of a last common ancestor but it does not include all of its descendants. The inclusion of the Carcharocles sharks in Otodus would make it monophyletic, with the sister clade being Megalolamna.[6]
The cladogram below represents the hypothetical relationships between megalodon and other sharks, including the great white shark. Modified from Shimada et al. (2016),[6] Ehret et al., (2009),[29] and the findings of Siversson et al. (2013).[38][39]
Biology
Appearance
One interpretation on how megalodon appeared was that it was a robust-looking shark, and may have had a similar build to the great white shark. The jaws may have been blunter and wider than the great white, and the fins would have also been similar in shape, though thicker due to its size. It may have had a pig-eyed appearance, in that it had small, deep-set eyes.[40]
Another interpretation is that megalodon bore a similarity to the whale shark (Rhincodon typus) or the basking shark (Cetorhinus maximus). The tail fin would have been crescent-shaped, the anal fin and second dorsal fin would have been small, and there would have been a caudal keel present on either side of the tail fin (on the caudal peduncle). This build is common in other large aquatic animals, such as whales, tuna, and other sharks, in order to reduce drag while swimming. The head shape can vary between species as most of the drag-reducing adaptations are toward the tail-end of the animal.[31]: 35–36
Sculpture in the Museum of Evolution in Puebla, Mexico
Since Carcharocles is derived from Otodus, and the two had teeth that bear a close similarity to those of the sand tiger shark (Carcharias taurus), megalodon may have had a build more similar to the sand tiger shark than to other sharks. This is unlikely since the sand tiger shark is a carangiform swimmer which requires faster movement of the tail for propulsion through the water than the great white shark, a thunniform swimmer.[31]: 35–36 [41]
Size
Due to fragmentary remains, there have been many contradictory size estimates for megalodon, as they can only be drawn from fossil teeth and vertebrae.[42]: 87 [43] The great white shark has been the basis of reconstruction and size estimation, as it is regarded as the best analogue to megalodon. Several total length estimation methods have been produced from comparing megalodon teeth and vertebrae to those of the great white.[40][44][10][7]
Size comparison of the great white and whale shark to estimates for megalodon
Proportions of megalodon at lengths of 3 m (10 ft), 8 m (26 ft), and 16 m (52 ft), extrapolated from extant relatives, with a 1.65 m (5 ft 5 in) diver
Megalodon size estimates vary depending on the method used, with maximum total length estimates ranging from 14.2–20.3 meters (47–67 ft).[40][10][7] A 2015 study estimated the average total body length at 10.5 meters (34 ft), calculated from 544 megalodon teeth, found throughout geological time and geography, including adults and juveniles.[11][12] In comparison, large great white sharks are generally around 6 meters (20 ft) in length, with a few contentious reports suggesting larger sizes.[45][46][40] The whale shark is the largest living fish, with one large female reported with a precaudal length of 15 meters (49 ft) and an estimated total length of 18.8 meters (62 ft).[45][47] It is possible that different populations of megalodon around the globe had different body sizes and behaviors due to different ecological pressures.[12] Megalodon is thought to have been the largest macropredatory shark that ever lived.[40]
«A C. megalodon about 16 meters long would have weighed about 48 metric tons (53 tons). A 17-meter (56-foot) C. megalodon would have weighed about 59 metric tons (65 tons), and a 20.3-meter (67 foot) monster would have topped off at 103 metric tons (114 tons).»[13]
In his 2015 book, The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution, Donald Prothero proposed the body mass estimates for different individuals of different length by extrapolating from a vertebral centra based on the dimensions of the great white,[13] a methodology also used for the 2008 study which supports the maximum mass estimate.[14]
In 2020, Cooper and his colleagues reconstructed a 2D model of megalodon based on the dimensions of all the extant lamnid sharks and suggested that a 16 meters (52 ft) long megalodon would have had a 4.65 m (15.3 ft) long head, 1.41 m (4 ft 8 in) tall gill slits, a 1.62 m (5 ft 4 in) tall dorsal fin, 3.08 m (10 ft 1 in) long pectoral fins, and a 3.85 m (12 ft 8 in) tall tail fin.[8] In 2022, Cooper and his colleagues also reconstructed a 3D model with the same basis as the 2020 study, resulting in a body mass estimate of 61.56 metric tons (67.86 short tons) for a 16 meters (52 ft) long megalodon (higher than the previous estimates); a vertebral column specimen named IRSNB P 9893 (formerly IRSNB 3121), belonging to a 46 year old individual from Belgium, was used for extrapolation. An individual of this size would have required 98,175 kcal per day, 20 times more than what the adult great white requires.[15]
Mature male megalodon may have had a body mass of 12.6 to 33.9 metric tons (13.9 to 37.4 short tons), and mature females may have been 27.4 to 59.4 metric tons (30.2 to 65.5 short tons), assuming that males could range in length from 10.5 to 14.3 meters (34 to 47 ft) and females 13.3 to 17 meters (44 to 56 ft).[40]
A 2015 study linking shark size and typical swimming speed estimated that megalodon would have typically swum at 18 kilometers per hour (11 mph)–assuming that its body mass was typically 48 metric tons (53 short tons)–which is consistent with other aquatic creatures of its size, such as the fin whale (Balaenoptera physalus) which typically cruises at speeds of 14.5 to 21.5 km/h (9.0 to 13.4 mph).[48] In 2022, Cooper and his colleagues converted this calculation into relative cruising speed (body lengths per second), resulting in an mean absolute cruising speed of 5 kilometers per hour (3.1 mph) and a mean relative cruising speed of 0.09 body lengths per second for a 16 meters (52 ft) long megalodon; the authors found their mean absolute cruising speed to be faster than any extant lamnid sharks and their mean relative cruising speed to be slower, consistent with previous estiamtes.[15]
Its large size may have been due to climatic factors and the abundance of large prey items, and it may have also been influenced by the evolution of regional endothermy (mesothermy) which would have increased its metabolic rate and swimming speed. The otodontid sharks have been considered to have been ectotherms, so on that basis megalodon would have been ectothermic. However, the largest contemporary ectothermic sharks, such as the whale shark, are filter feeders, while lamnids are now known to be regional endotherms, implying some metabolic correlations with a predatory lifestyle. These considerations, as well as tooth oxygen isotopic data and the need for higher burst swimming speeds in macropredators of endothermic prey than ectothermy would allow, imply that otodontids, including megalodon, were probably regional endotherms.[49]
In 2020, Shimada and colleagues suggested large size was instead due to intrauterine cannibalism, where the larger fetus eats the smaller fetus, resulting in progressively larger and larger fetuses, requiring the mother to attain even greater size as well as caloric requirements which would have promoted endothermy. Males would have needed to keep up with female size in order to still effectively copulate (which probably involved latching onto the female with claspers, like modern cartilaginous fish).[50]
Maximum estimates
The first attempt to reconstruct the jaw of megalodon was made by Bashford Dean in 1909, displayed at the American Museum of Natural History. From the dimensions of this jaw reconstruction, it was hypothesized that megalodon could have approached 30 meters (98 ft) in length. Dean had overestimated the size of the cartilage on both jaws, causing it to be too tall.[51][52]
Tooth compared to hand
In 1973, John E. Randall, an ichthyologist, used the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) to measure the length of the shark, yielding a maximum length of about 13 meters (43 ft).[53] However, tooth enamel height does not necessarily increase in proportion to the animal’s total length.[30]: 99
In 1994, marine biologists Patrick J. Schembri and Stephen Papson opined that O. megalodon may have approached a maximum of around 24 to 25 meters (79 to 82 ft) in total length.[54][55]
In 1996, shark researchers Michael D. Gottfried, Leonard Compagno, and S. Curtis Bowman proposed a linear relationship between the great white shark’s total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in meters = − (0.096) × [UA maximum height (mm)]-(0.22).[56][40] Using this tooth height regression equation, the authors estimated a total length of 15.9 meters (52 ft) based on a tooth 16.8 centimeters (6.6 in) tall, which the authors considered a conservative maximum estimate. They also compared the ratio between the tooth height and total length of large female great whites to the largest megalodon tooth. A 6-meter (20 ft) long female great white, which the authors considered the largest ‘reasonably trustworthy’ total length, produced an estimate of 16.8 meters (55 ft). However, based on the largest female great white reported, at 7.1 meters (23 ft), they estimated a maximum estimate of 20.2 meters (66 ft).[40]
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimeter (0.39 in) of root width, there are approximately 1.4 meters (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah’s possession had a root width of about 12 centimeters (4.7 in), which yielded 16.5 meters (54 ft) in total length.[31]: 88
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length after conducting anatomical analysis of several specimens, allowing any sized tooth to be used. Shimada stated that the previously proposed methods were based on a less-reliable evaluation of the dental homology between megalodon and the great white shark, and that the growth rate between the crown and root is not isometric, which he considered in his model. Using this model, the upper anterior tooth possessed by Gottfried and colleagues corresponded to a total length of 15 meters (49 ft).[57] Among several specimens found in the Gatún Formation of Panama, one upper lateral tooth was used by other researchers to obtain a total length estimate of 17.9 meters (59 ft) using this method.[36][58]
In 2019, Shimada revisited the size of megalodon and discouraged using non-anterior teeth for estimations, noting that the exact position of isolated non-anterior teeth is difficult to identify. Shimada provided maximum total length estimates using the largest anterior teeth available in museums. The tooth with the tallest crown height known to Shimada, NSM PV-19896, produced a total length estimate of 14.2 meters (47 ft). The tooth with the tallest total height, FMNH PF 11306, was reported at 16.8 centimeters (6.6 in). However, Shimada remeasured the tooth and found it actually to measure 16.2 centimeters (6.4 in). Using the total height tooth regression equation proposed by Gottfried and colleagues produced an estimate of 15.3 meters (50 ft).[7][10]
In 2021, Victor J. Perez, Ronny M. Leder, and Teddy Badaut proposed a method of estimating total length of megalodon from the sum of the tooth crown widths. Using more complete megalodon dentitions, they reconstructed the dental formula and then made comparisons to living sharks. The researchers noted that the 2002 Shimada crown height equations produce wildly varying results for different teeth belonging to the same shark (range of error of ± 9 metres (30 ft)), casting doubt on some of the conclusions of previous studies using that method. Using the largest tooth available to the authors, GHC 6, with a crown width of 13.3 centimeters (5.2 in), they estimated a maximum body length of approximately 20 meters (66 ft), with a range of error of approximately ± 3.5 metres (11 ft).[10] This maximum length estimate was also supported by Cooper and his colleagues in 2022.[15]
There are anecdotal reports of teeth larger than those found in museum collections.[7] Gordon Hubbell from Gainesville, Florida, possesses an upper anterior megalodon tooth whose maximum height is 18.4 centimeters (7.25 in), one of the largest known tooth specimens from the shark.[59] In addition, a 2.7-by-3.4-meter (9 by 11 ft) megalodon jaw reconstruction developed by fossil hunter Vito Bertucci contains a tooth whose maximum height is reportedly over 18 centimeters (7 in).[60]
Teeth and bite force
The most common fossils of megalodon are its teeth. Diagnostic characteristics include a triangular shape, robust structure, large size, fine serrations, a lack of lateral denticles, and a visible V-shaped neck (where the root meets the crown).[30]: 55 [36] The tooth met the jaw at a steep angle, similar to the great white shark. The tooth was anchored by connective tissue fibers, and the roughness of the base may have added to mechanical strength.[61] The lingual side of the tooth, the part facing the tongue, was convex; and the labial side, the other side of the tooth, was slightly convex or flat. The anterior teeth were almost perpendicular to the jaw and symmetrical, whereas the posterior teeth were slanted and asymmetrical.[62]
Megalodon teeth can measure over 180 millimeters (7.1 in) in slant height (diagonal length) and are the largest of any known shark species,[31]: 33 implying it was the largest of all macropredatory sharks.[40] In 1989, a nearly complete set of megalodon teeth was discovered in Saitama, Japan. Another nearly complete associated megalodon dentition was excavated from the Yorktown Formations in the United States, and served as the basis of a jaw reconstruction of megalodon at the National Museum of Natural History (USNM). Based on these discoveries, an artificial dental formula was put together for megalodon in 1996.[30]: 55 [63]
The dental formula of megalodon is: 2.1.7.43.0.8.4. As evident from the formula, megalodon had four kinds of teeth in its jaws: anterior, intermediate, lateral, and posterior. Megalodon’s intermediate tooth technically appears to be an upper anterior and is termed as «A3» because it is fairly symmetrical and does not point mesially (side of the tooth toward the midline of the jaws where the left and right jaws meet). Megalodon had a very robust dentition,[30]: 20–21 and had over 250 teeth in its jaws, spanning 5 rows.[31]: iv It is possible that large megalodon individuals had jaws spanning roughly 2 meters (6.6 ft) across.[31]: 129 The teeth were also serrated, which would have improved efficiency in cutting through flesh or bone.[19][31]: 1 The shark may have been able to open its mouth to a 75° angle, though a reconstruction at the USNM approximates a 100° angle.[40]
In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5-meter (8.2 ft) long specimen, and then isometrically scaled the results for its maximum size and the conservative minimum and maximum body mass of megalodon. They placed the bite force of the latter between 108,514 to 182,201 newtons (24,395 to 40,960 lbf) in a posterior bite, compared to the 18,216 newtons (4,095 lbf) bite force for the largest confirmed great white shark, and 7,400 newtons (1,700 lbf) for the placoderm fish Dunkleosteus. In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the force generated, which would probably have caused the total force experienced by prey to be higher than the estimate.[14][64]
In 2021, Antonio Ballell and Humberto Ferrón used Finite Element Analysis modeling to examine the stress distribution of three types of megalodon teeth and closely related mega-toothed species when exposed to anterior and lateral forces, the latter of which would be generated when a shark shakes its head to tear through flesh. The resulting simulations identified higher levels of stress in megalodon teeth under lateral force loads compared to its precursor species such as O. obliquus and O. angusteidens when tooth size was removed as a factor. This suggests that megalodon teeth were of a different functional significance than previously expected, challenging prior interpretations that megalodon’s dental morphology was primarily driven by a dietary shift towards marine mammals. Instead, the authors proposed that it was a byproduct of an increase in body size caused by heterochronic selection.[65]
Internal anatomy
Megalodon is represented in the fossil record by teeth, vertebral centra, and coprolites.[40][66] As with all sharks, the skeleton of megalodon was formed of cartilage rather than bone; consequently most fossil specimens are poorly preserved.[67] To support its large dentition, the jaws of megalodon would have been more massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. Its chondrocranium, the cartilaginous skull, would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size.[40]
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp Basin, Belgium, in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimeters (2.2 in) to 155 millimeters (6 in) in diameter. The shark’s vertebrae may have gotten much bigger, and scrutiny of the specimen revealed that it had a higher vertebral count than specimens of any known shark, possibly over 200 centra; only the great white approached it.[40] Another partially preserved vertebral column of a megalodon was excavated from the Gram Formation in Denmark in 1983, which comprises 20 vertebral centra, with the centra ranging from 100 millimeters (4 in) to 230 millimeters (9 in) in diameter.[61]
The coprolite remains of megalodon are spiral-shaped, indicating that the shark may have had a spiral valve, a corkscrew-shaped portion of the lower intestines, similar to extant lamniform sharks. Miocene coprolite remains were discovered in Beaufort County, South Carolina, with one measuring 14 cm (5.5 in).[66]
Gottfried and colleagues reconstructed the entire skeleton of megalodon, which was later put on display at the Calvert Marine Museum in the United States and the Iziko South African Museum.[40][32] This reconstruction is 11.3 meters (37 ft) long and represents a mature male,[40]: 61 based on the ontogenetic changes a great white shark experiences over the course of its life.[40]: 65
Paleobiology
Range and habitat
Megalodon had a cosmopolitan distribution;[26][58] its fossils have been excavated from many parts of the world, including Europe, Africa, the Americas, and Australia.[30]: 67 [68] It most commonly occurred in subtropical to temperate latitudes.[26][30]: 78 It has been found at latitudes up to 55° N; its inferred tolerated temperature range was 1–24 °C (34–75 °F).[note 1] It arguably had the capacity to endure such low temperatures due to mesothermy, the physiological capability of large sharks to maintain a higher body temperature than the surrounding water by conserving metabolic heat.[26]
Megalodon inhabited a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult megalodon were not abundant in shallow water environments, and mostly inhabited offshore areas. Megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.[31]: 33 [70]
Fossil remains show a trend for specimens to be larger on average in the Southern Hemisphere than in the Northern, with mean lengths of 11.6 and 9.6 meters (38 and 31 ft), respectively; and also larger in the Pacific than the Atlantic, with mean lengths of 10.9 and 9.5 meters (36 and 31 ft) respectively. They do not suggest any trend of changing body size with absolute latitude, or of change in size over time (although the Carcharocles lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 meters (34 ft), with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.[12]
Locations of fossils
Megalodon had a global distribution and fossils of the shark have been found in many places around the world, bordering all oceans of the Neogene.[71]
| Epoch | Formation | State | Continent |
|---|---|---|---|
| Pliocene | Luanda Formation | Africa | |
| Africa | |||
| Africa | |||
| Castell’Arquato Formation | Europe | ||
| Arenas de Huelva Formation | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Touril Complex Formation | Europe | ||
| Red Crag Formation | Europe | ||
| San Mateo Formation | North America | ||
| Towsley Formation | North America | ||
| Bone Valley Formation | North America | ||
| Tamiami Formation | North America | ||
| Yorktown Formation | North America | ||
| Highlands Formation | North America | ||
| Refugio Formation | North America | ||
| San Diego Formation | North America | ||
| Tirabuzon Formation | North America | ||
| Onzole Formation | South America | ||
| Paraguaná Formation | South America | ||
| Black Rock Sandstone | Oceania | ||
| Cameron Inlet Formation | Oceania | ||
| Grange Burn Formation | Oceania | ||
| Loxton Sand Formation | Oceania | ||
| Whaler’s Bluff Formation | Oceania | ||
| Tangahoe Formation | Oceania | ||
| Miocene | |||
| Africa | |||
| Madagascar Basin | Africa | ||
| Africa | |||
| Varswater Formation | Africa | ||
| Baripada Limestone | Asia | ||
| Arakida Formation | Asia | ||
| Bihoku Group | Asia | ||
| Fujina Formation | Asia | ||
| Hannoura Formation | Asia | ||
| Hongo Formation | Asia | ||
| Horimatsu Formation | Asia | ||
| Ichishi Formation | Asia | ||
| Kurahara Formation | Asia | ||
| Maenami Formation | Asia | ||
| Matsuyama Group | Asia | ||
| Sekinobana Formation | Asia | ||
| Suso Formation | Asia | ||
| Takakubo Formation | Asia | ||
| Tonokita Formation | Asia | ||
| Tsurushi Formation | Asia | ||
| Wajimazaki Formation | Asia | ||
| Yoshii Formation | Asia | ||
| Asia | |||
| Burgeschleinitz Formation | Europe | ||
| Melker Sand Formation | Europe | ||
| Rzehakia Formation | Europe | ||
| Weissenegg Formation | Europe | ||
| Antwerpen Sands Member | Europe | ||
| Europe | |||
| Hrušky Formation | Europe | ||
| Gram Formation | Europe | ||
| Aquitaine Basin | Europe | ||
| Europe | |||
| Libano Sandstone | Europe | ||
| Blue Clay Formation | Europe | ||
| Globigerina Limestone[72] | Europe | ||
| Aalten Member | Europe | ||
| Breda Formation | Europe | ||
| Korytnica Clays | Europe | ||
| Leitha Limestone | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Filakovo Formation | Europe | ||
| Arjona Formation | Europe | ||
| Calcarenites of Sant Elm | Europe | ||
| Europe | |||
| Monterey Formation | North America | ||
| Puente Formation | North America | ||
| Purisima Formation | North America | ||
| San Mateo Formation | North America | ||
| Santa Margarita Formation | North America | ||
| Temblor Formation | North America | ||
| Topanga Formation | North America | ||
| Bone Valley Formation | North America | ||
| Calvert Formation | North America | ||
| Kirkwood Formation | North America | ||
| North America | |||
| Cojímar Formation | North America | ||
| Kendance Formation | North America | ||
| North America | |||
| Aymamón Limestone | North America | ||
| Almejas Formation | North America | ||
| Carrillo Puerto Formation | North America | ||
| Chagres Formation | North America | ||
| Chucunaque Formation | North America | ||
| Gatún Formation | North America | ||
| Paraná Formation | South America | ||
| Bahía Inglesa Formation | South America | ||
| Castilletes Formation | South America | ||
| Miramar Formation | South America | ||
| Pisco Formation | South America | ||
| Camacho Formation | South America | ||
| Cantaure Formation | South America | ||
| Caujarao Formation | South America | ||
| Socorro Formation | South America | ||
| Urumaco Formation | South America | ||
| Batesford Limestone | Oceania | ||
| Black Rock Sandstone | Oceania | ||
| Gippsland Limestone | Oceania | ||
| Mannum Formation | Oceania | ||
| Morgan Limestone | Oceania | ||
| Port Campbell Limestone | Oceania | ||
| Oceania | |||
| Oceania | |||
| Duho Formation[73] | Asia | ||
| Seogwipo Formation[74] | Asia |
Locations of megalodon fossil discoveries, yellow from the Pliocene and blue from the Miocene[26][71]
Prey relationships
Vertebra of a whale bitten in half by a megalodon with visible gashes from teeth.
Though sharks are generally opportunistic feeders, megalodon’s great size, high-speed swimming capability, and powerful jaws, coupled with an impressive feeding apparatus, made it an apex predator capable of consuming a broad spectrum of animals. Otodus megalodon was probably one of the most powerful predators to have existed.[15] A study focusing on calcium isotopes of extinct and extant elasmobranch sharks and rays revealed that megalodon fed at a higher trophic level than the contemporaneous great white shark («higher up» in the food chain.)[75]
Fossil evidence indicates that megalodon preyed upon many cetacean species, such as dolphins, small whales, cetotheres, squalodontids (shark toothed dolphins), sperm whales, bowhead whales, and rorquals.[51][76][77] In addition to this, they also targeted seals, sirenians, and sea turtles.[70] The shark was an opportunist and piscivorous, and it would have also gone after smaller fish and other sharks.[51] Many whale bones have been found with deep gashes most likely made by their teeth.[30]: 75 Various excavations have revealed megalodon teeth lying close to the chewed remains of whales,[30]: 75 [32] and sometimes in direct association with them.[78]
The feeding ecology of megalodon appears to have varied with age and between sites, like the modern great white shark. It is plausible that the adult megalodon population off the coast of Peru targeted primarily cetothere whales 2.5 to 7 meters (8.2 to 23 ft) in length and other prey smaller than itself, rather than large whales in the same size class as themselves.[76] Meanwhile, juveniles likely had a diet that consisted more of fish.[36][79]
Competition
Megalodon faced a highly competitive environment.[80] Its position at the top of the food chain[81] probably had a significant impact on the structuring of marine communities.[80][82] Fossil evidence indicates a correlation between megalodon and the emergence and diversification of cetaceans and other marine mammals.[30]: 78 [80] Juvenile megalodon preferred habitats where small cetaceans were abundant, and adult megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.[30]: 74–75
Megalodon were contemporaneous with whale-eating toothed whales (particularly macroraptorial sperm whales and squalodontidae), which were also probably among the era’s apex predators, and provided competition.[80] Some attained gigantic sizes, such as Livyatan, estimated between 13.5 to 17.5 meters (44 to 57 ft). Fossilized teeth of an undetermined species of such physeteroids from Lee Creek Mine, North Carolina, indicate it had a maximum body length of 8–10 m and a maximum lifespan of about 25 years. This is very different from similarly sized modern killer whales that live to 65 years, suggesting that unlike the latter, which are apex predators, these physeteroids were subject to predation from larger species such as megalodon or Livyatan.[83] By the Late Miocene, around 11 Mya, macroraptorials experienced a significant decline in abundance and diversity. Other species may have filled this niche in the Pliocene,[80][84] such as the fossil killer whale Orcinus citoniensis which may have been a pack predator and targeted prey larger than itself,[32][85][86][87] but this inference is disputed,[28] and it was probably a generalist predator rather than a marine mammal specialist.[88]
Megalodon may have subjected contemporaneous white sharks to competitive exclusion, as the fossil records indicate that other shark species avoided regions it inhabited by mainly keeping to the colder waters of the time.[89][30]: 77 In areas where their ranges seemed to have overlapped, such as in Pliocene Baja California, it is possible that megalodon and the great white shark occupied the area at different times of the year while following different migratory prey.[30]: 77 [90] Megalodon probably also had a tendency for cannibalism, much like contemporary sharks.[91]
Feeding strategies
Sharks often employ complex hunting strategies to engage large prey animals. Great white shark hunting strategies may be similar to how megalodon hunted its large prey.[92] Megalodon bite marks on whale fossils suggest that it employed different hunting strategies against large prey than the great white shark.[51]
One particular specimen–the remains of a 9-meter (30 ft) long undescribed Miocene baleen whale–provided the first opportunity to quantitatively analyze its attack behavior. Unlike great whites which target the underbelly of their prey, megalodon probably targeted the heart and lungs, with their thick teeth adapted for biting through tough bone, as indicated by bite marks inflicted to the rib cage and other tough bony areas on whale remains.[51] Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans, for example cetotheres, suggest that they were rammed with great force from below before being killed and eaten, based on compression fractures.[92]
There is also evidence that a possible separate hunting strategy existed for attacking raptorial sperm whales; a tooth belonging to an undetermined 4 m (13 ft) physeteroid closely resembling those of Acrophyseter discovered in the Nutrien Aurora Phosphate Mine in North Carolina suggests that a megalodon or O. chubutensis may have aimed for the head of the sperm whale in order to inflict a fatal bite, the resulting attack leaving distinctive bite marks on the tooth. While scavenging behavior cannot be ruled out as a possibility, the placement of the bite marks is more consistent with predatory attacks than feeding by scavenging, as the jaw is not a particularly nutritious area to for a shark feed or focus on. The fact that the bite marks were found on the tooth’s roots further suggest that the shark broke the whale’s jaw during the bite, suggesting the bite was extremely powerful. The fossil is also notable as it stands as the first known instance of an antagonistic interaction between a sperm whale and an otodontid shark recorded in the fossil record.[93]
During the Pliocene, larger cetaceans appeared.[94] Megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones and tail vertebrae of large whales from the Pliocene have been found with megalodon bite marks, which suggests that megalodon would immobilize a large whale before killing and feeding on it.[14][51]
Growth and reproduction
In 2010, Ehret estimated that megalodon had a fast growth rate nearly two times that of the extant great white shark. He also estimated that the slowing or cessation of somatic growth in megalodon occurred around 25 years of age, suggesting that this species had an extremely delayed sexual maturity.[95] In 2021, Shimada and colleagues calculated the growth rate of an approximately 9.2 m (30 ft) individual based on the Belgian vertebrate column specimen that presumably contains annual growth rings on three of its vertebrae. They estimated the individual died at 46 years of age, with a growth rate of 16 cm (6.3 in) per year, and a length of 2 m (6 ft 7 in) at birth. For a 15 m (49 ft) individual—which they considered to have been the maximum size attainable—this would equate to a lifespan of 88 to 100 years.[96] However, Cooper and his colleagues in 2022 estimated the length of this 46 year old individual at nearly 16 m (52 ft) based on the 3D reconstruction which resulted in the complete vertebral column to be 11.1 m (36 ft) long; the researchers claimed that this size estimate difference occurred due to the fact that Shimada and his colleagues extrapolated its size only based on the vertebral centra.[15]
Megalodon, like contemporaneous sharks, made use of nursery areas to birth their young in, specifically warm-water coastal environments with large amounts of food and protection from predators.[36] Nursery sites were identified in the Gatún Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands,[97] and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of megalodon also.[98] Infant megalodons were around 3.5 meters (11 ft) at their smallest,[40]: 61 and the pups were vulnerable to predation by other shark species, such as the great hammerhead shark (Sphyrna mokarran) and the snaggletooth shark (Hemipristis serra).[36] Their dietary preferences display an ontogenetic shift:[40]: 65 Young megalodon commonly preyed on fish,[36] sea turtles,[70] dugongs,[31]: 129 and small cetaceans; mature megalodon moved to off-shore areas and consumed large cetaceans.[30]: 74–75
An exceptional case in the fossil record suggests that juvenile megalodon may have occasionally attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-to-7-meter (13 to 23 ft) long Pliocene shark were found on a rib from an ancestral blue or humpback whale that showed evidence of subsequent healing, which is suspected to have been inflicted by a juvenile megalodon.[99][100]
Extinction
Climate change
The Earth experienced a number of changes during the time period megalodon existed which affected marine life. A cooling trend starting in the Oligocene 35 Mya ultimately led to glaciation at the poles. Geological events changed currents and precipitation; among these were the closure of the Central American Seaway and changes in the Tethys Ocean, contributing to the cooling of the oceans. The stalling of the Gulf Stream prevented nutrient-rich water from reaching major marine ecosystems, which may have negatively affected its food sources. The largest fluctuation of sea levels in the Cenozoic era occurred in the Plio-Pleistocene, between around 5 million to 12 thousand years ago, due to the expansion of glaciers at the poles, which negatively impacted coastal environments, and may have contributed to its extinction along with those of several other marine megafaunal species.[101] These oceanographic changes, in particular the sea level drops, may have restricted many of the suitable shallow warm-water nursery sites for megalodon, hindering reproduction.[102] Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.[103][36]
As its range did not apparently extend into colder waters, megalodon may not have been able to retain a significant amount of metabolic heat, so its range was restricted to shrinking warmer waters.[102][77][104] Fossil evidence confirms the absence of megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[30]: 77 However, an analysis of the distribution of megalodon over time suggests that temperature change did not play a direct role in its extinction. Its distribution during the Miocene and Pliocene did not correlate with warming and cooling trends; while abundance and distribution declined during the Pliocene, megalodon did show a capacity to inhabit colder latitudes. It was found in locations with a mean temperature ranging from 12 to 27 °C (54 to 81 °F), with a total range of 1 to 33 °C (34 to 91 °F), indicating that the global extent of suitable habitat should not have been greatly affected by the temperature changes that occurred.[26] This is consistent with evidence that it was a mesotherm.[49]
Changing ecosystem
Marine mammals attained their greatest diversity during the Miocene,[30]: 71 such as with baleen whales with over 20 recognized Miocene genera in comparison to only six extant genera.[106] Such diversity presented an ideal setting to support a super-predator such as megalodon.[30]: 75 By the end of the Miocene, many species of mysticetes had gone extinct;[80] surviving species may have been faster swimmers and thus more elusive prey.[31]: 46 Furthermore, after the closure of the Central American Seaway, tropical whales decreased in diversity and abundance.[104] The extinction of megalodon correlates with the decline of many small mysticete lineages, and it is possible that it was quite dependent on them as a food source.[76] Additionally, a marine megafauna extinction during the Pliocene was discovered to have eliminated 36% of all large marine species including 55% of marine mammals, 35% of seabirds, 9% of sharks, and 43% of sea turtles. The extinction was selective for endotherms and mesotherms relative to poikilotherms, implying causation by a decreased food supply[101] and thus consistent with megalodon being mesothermic.[49] Megalodon may have been too large to sustain itself on the declining marine food resources.[102] The cooling of the oceans during the Pliocene might have restricted the access of megalodon to the polar regions, depriving it of the large whales which had migrated there.[104]
Competition from large odontocetes, such as macropredatory sperm whales which appeared in the Miocene, and a member of genus Orcinus (i.e., Orcinus citoniensis) in the Pliocene,[80][84] is assumed to have contributed to the decline and extinction of megalodon.[26][31]: 46–47 [102][107] But this assumption is disputed:[28] The Orcininae emerged in Mid-Pliocene with O. citoniensis reported from the
Pliocene of Italy,[84][108] and similar forms reported from the Pliocene of England and South Africa,[84] indicating the capacity of these dolphins to cope with increasingly prevalent cold water temperatures in high latitudes.[84] These dolphins were assumed to have been macrophagous in some studies,[26] but on closer inspection, these dolphins are not found to be macrophagous and fed on small fishes instead.[108] On the other hand, gigantic macropredatory sperm whales such as Livyatan-like forms are last reported from Australia and South Africa circa 5 million years ago.[109][110][111] Others, such as Hoplocetus and Scaldicetus also occupied a niche similar to that of modern killer whales but the last of these forms disappeared during the Pliocene.[112][108] Members of genus Orcinus became large and macrophagous in the Pleistocene.[108]
Paleontologist Robert Boessenecker and his colleagues rechecked the fossil record of megalodon for carbon dating errors and concluded that it disappeared circa 3.5 million years ago.[28] Boessenecker and his colleagues further suggest that megalodon suffered range fragmentation due to climatic shifts,[28] and competition with white sharks might have contributed to its decline and extinction.[28] Competition with white sharks is assumed to be a factor in other studies as well,[113][26][107] but this hypothesis warrants further testing.[114] Multiple compounding environmental and ecological factors including climate change and thermal limitations, collapse of prey populations and resource competition with white sharks are believed to have contributed to decline and extinction of megalodon for now.[107]
The extinction of megalodon set the stage for further changes in marine communities. The average body size of baleen whales increased significantly after its disappearance, although possibly due to other, climate-related, causes.[115] Conversely the increase in baleen whale size may have contributed to the extinction of megalodon, as they may have preferred to go after smaller whales; bite marks on large whale species may have come from scavenging sharks. Megalodon may have simply become coextinct with smaller whale species, such as Piscobalaena nana.[105] The extinction of megalodon had a positive impact on other apex predators of the time, such as the great white shark, in some cases spreading to regions where megalodon became absent.[26][113][116]
In popular culture
HMS Challenger discovered megalodon teeth which were erroneously dated to be around 11,000 to 24,000 years old.
Megalodon has been portrayed in many works of fiction, including films and novels, and continues to be a popular subject for fiction involving sea monsters.[117] Reports of supposedly fresh megalodon teeth, such as those found by HMS Challenger in 1873 which were dated in 1959 by the zoologist Wladimir Tschernezky to be around 11,000 to 24,000 years old, helped popularise claims of recent megalodon survival amongst cryptozoologists.[118] These claims are now discredited, and are probably teeth that were well-preserved by a thick mineral-crust precipitate of manganese dioxide, and so had a lower decomposition rate and retained a white color during fossilization. Fossil megalodon teeth can vary in color from off-white to dark browns and greys, and some fossil teeth may have been redeposited into a younger stratum. The claims that megalodon could remain elusive in the depths, similar to the megamouth shark which was discovered in 1976, are unlikely as the shark lived in warm coastal waters and probably could not survive in the cold and nutrient-poor deep sea environment.[119][120]
Contemporary fiction about megalodon surviving into modern times was pioneered by the 1997 novel Meg: A Novel of Deep Terror by Steve Alten and its subsequent sequels. Megalodon subsequently began to feature in films, such as the 2003 direct to video Shark Attack 3: Megalodon, and later The Meg, a 2018 film based on the 1997 book which grossed over $500 million at the box office.[118][121]
Animal Planet’s pseudo-documentary Mermaids: The Body Found included an encounter 1.6 mya between a pod of mermaids and a megalodon.[122] Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives,[123] a controversial docufiction about the creature that presented alleged evidence in order to suggest that megalodons still lived. This program received criticism for being completely fictional and for inadequately disclosing its fictional nature; for example, all of the supposed scientists depicted were paid actors, and there was no disclosure in the documentary itself that it was fictional. In a poll by Discovery, 73% of the viewers of the documentary thought that megalodon was not extinct. In 2014, Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.[51][124][125][126] Despite the criticism from scientists, Megalodon: The Monster Shark Lives was a huge ratings success, gaining 4.8 million viewers, the most for any Shark Week episode up to that point.[127]
Megalodon teeth are the state fossil of North Carolina.[128]
See also
- List of prehistoric cartilaginous fish
- Prehistoric fish
- Largest prehistoric organisms
Notes
- ^ Carbonated bioapatite from a megalodon tooth (of unknown source location) dated to 5.75 ± 0.9 Ma in age has been analyzed for isotope ratios of oxygen (18O/16O) and carbon (13C/12C), using a carbonate clumped-isotope thermometer methodology to yield an estimate of the ambient temperature in that individual’s environment of 19 ± 4 °C.[69]
References
- ^ a b Agassiz, Louis (1843). Recherches sur les poissons fossiles [Research on the fossil fishes] (in French). Neuchatel: Petitpierre. p. 41.
- ^ «Otodus (Megaselachus) megalodon (Agassiz, 1837)». SharkReferences.com. Retrieved 24 October 2017.
- ^ Eastman, C. R. (1904). Maryland Geological Survey. Vol. 2. Baltimore, Maryland: Johns Hopkins University. p. 82.
- ^ a b c Cappetta, H. (1987). «Mesozoic and Cenozoic Elasmobranchii». Handbook of Paleoichthyology. Vol. 3B. München, Germany: Friedrich Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- ^ Hay, O. P. (1901). «Bibliography and Catalogue of the Fossil Vertebrata of North America». Bulletin of the United States Geological Society (179): 308.
- ^ a b c d e f Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (2016). «A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade». Historical Biology. 29 (5): 1–11. doi:10.1080/08912963.2016.1236795. S2CID 89080495.
- ^ a b c d e f Shimada, Kenshu (2019). «The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited». Historical Biology. 33 (7): 1–8. doi:10.1080/08912963.2019.1666840. ISSN 0891-2963. S2CID 208570844.
- ^ a b Cooper, J. A.; Pimiento, C.; Ferrón, H. G.; Benton, M. J. (2020). «Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction». Scientific Reports. 10 (14596): 14596. Bibcode:2020NatSR..1014596C. doi:10.1038/s41598-020-71387-y. PMC 7471939. PMID 32883981.
- ^ «Giant ‘megalodon’ shark extinct earlier than previously thought». Science Daily. 13 February 2019.
- ^ a b c d e Perez, Victor; Leder, Ronny; Badaut, Teddy (2021). «Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions». Palaeontologia Electronica. 24 (1): 1–28. doi:10.26879/1140.
- ^ a b Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c d e Pimiento, C.; Balk, M. A. (2015). «Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators». Paleobiology. 41 (3): 479–490. doi:10.1017/pab.2015.16. PMC 4541548. PMID 26321775.
- ^ a b c Prothero, Donald R. (25 August 2015). «09. Mega-Jaws: The Largest Fish. Carcharocles». The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution. New York Chichester, West Sussex: Columbia University Press. pp. 96–110. doi:10.7312/prot17190-010. ISBN 978-0-231-53942-5.
- ^ a b c d e Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?» (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ^ a b c d e f Cooper, J.A.; Hutchinson, J.R.; Bernvi, D.C.; Cliff, G.; Wilson, R.P.; Dicken, M.L.; Menzel, J.; Wroe, S.; Pirlo, J.; Pimiento, C. (2022). «The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling». Science Advances. 8 (33): eabm9424. Bibcode:2022SciA….8M9424C. doi:10.1126/sciadv.abm9424. ISSN 2375-2548. PMC 9385135. PMID 35977007.
- ^ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Westport, Connecticut: Libraries Unlimited. pp. 25–26. ISBN 978-1-59158-265-6. OCLC 230807846.
- ^ Hsu, Kuang-Tai (2009). «The Path to Steno’s Synthesis on the Animal Origin of Glossopetrae». In Rosenburg, G. D. (ed.). The Revolution in Geology from the Renaissance to the Enlightenment. Vol. 203. Boulder, Colorado: Geological Society of America. ISBN 978-0-8137-1203-1. OCLC 608657795.
- ^ Eilperin, J. (2012). Demon Fish. Pantheon Books. p. 43. ISBN 978-0-7156-4352-5.
- ^ a b c d e f Nyberg, K. G.; Ciampaglio C. N.; Wray G. A. (2006). «Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth». Journal of Vertebrate Paleontology. 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. S2CID 53640614.
- ^ Keyes, I. W. (2012). «New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record». New Zealand Journal of Geology and Geophysics. 15 (2): 229. doi:10.1080/00288306.1972.10421956.
- ^ μέγας. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ ὀδούς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ Lawley, R. (1881). «Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)» [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (in Italian). 5: 167–172.
- ^ Yabe, H.; Goto, M.; Kaneko, N. (2004). «Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records». The Palaeontological Society of Japan. 75: 7–15.
- ^ a b c Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ a b c d e f g h i j k l Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c Pimiento, C.; Clements, C. F. (2014). «When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record». PLOS ONE. 9 (10): e111086. Bibcode:2014PLoSO…9k1086P. doi:10.1371/journal.pone.0111086. PMC 4206505. PMID 25338197.
- ^ a b c d e f Boessenecker, R. W.; Ehret, D. J.; Long, D. J.; Churchill, M.; Martin, E.; Boessenecker, S. J. (2019). «The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific». PeerJ. 7: e6088. doi:10.7717/peerj.6088. PMC 6377595. PMID 30783558.
- ^ a b c d Ehret D. J.; Hubbell G.; Macfadden B. J. (2009). «Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru». Journal of Vertebrate Paleontology. 29 (1): 1–13. doi:10.1671/039.029.0113. JSTOR 20491064. S2CID 129585445.
- ^ a b c d e f g h i j k l m n o p q r Klimley, Peter; Ainley, David (1996). «Evolution». Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. ISBN 978-0-12-415031-7. OCLC 212425118.
- ^ a b c d e f g h i j k l m n o p Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ a b c d e Andres, Lutz (2002). «C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles«. fossilguy.com. Retrieved 16 January 2008.
- ^ a b Perez, V. J.; Godfrey, S. J.; Kent, B. W.; Weems, R. E.; Nance, J. R. (2019). «The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time». Journal of Vertebrate Paleontology. 38 (6): e1546732. doi:10.1080/02724634.2018.1546732.
- ^ Siverson, M.; Lindgren, J.; Newbrey, M.G.; Cederström, P.; Cook, T.D. (2013). «Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type» (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. S2CID 58906204. Archived from the original (PDF) on 19 October 2013.
- ^ Benton, M. J.; Pearson, P. N. (2001). «Speciation in the fossil record». Trends in Ecology and Evolution. 16 (7): 405–411. doi:10.1016/s0169-5347(01)02149-8. PMID 11403874.
- ^ a b c d e f g h Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Vivian, G. (2013). «Research debunks Great White lineage».
- ^ «Palaeontological detective work unravels evolution of megatooth sharks». 2013.
- ^ a b c d e f g h i j k l m n o p q r Gottfried, MD; Compagno, LJV; Bowman, SC (1996). «Size and skeletal anatomy of the giant megatooth shark Carcharodon megalodon«. In Klimley; Ainley (eds.). Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. pp. 55–89. ISBN 978-0124150317.
- ^ «Could Megalodon Have Looked Like a BIG Sandtiger Shark?». Biology of Sharks and Rays. Retrieved 2 September 2017.
- ^ Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). «Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles» (PDF). Caribbean Journal of Science. 44 (3): 279–286. doi:10.18475/cjos.v44i3.a2. S2CID 87154947. Archived from the original (PDF) on 20 July 2011.
- ^ Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ a b McClain, Craig R.; Balk, Meghan A.; Benfield, Mark C.; Branch, Trevor A.; Chen, Catherine; Cosgrove, James; Dove, Alistair D.M.; Gaskins, Lindsay C.; Helm, Rebecca R.; Hochberg, Frederick G.; Lee, Frank B. (13 January 2015). «Sizing ocean giants: patterns of intraspecific size variation in marine megafauna». PeerJ. 3: e715. doi:10.7717/peerj.715. ISSN 2167-8359. PMC 4304853. PMID 25649000.
- ^ I., Castro, Jose (2011). Sharks of North America. Oxford University Press, USA. ISBN 978-0-19-978097-6. OCLC 958576172.
- ^ Borrell, Asunción; Aguilar, Alex; Gazo, Manel; Kumarran, R. P.; Cardona, Luis (1 December 2011). «Stable isotope profiles in whale shark (Rhincodon typus) suggest segregation and dissimilarities in the diet depending on sex and size». Environmental Biology of Fishes. 92 (4): 559–567. doi:10.1007/s10641-011-9879-y. ISSN 1573-5133. S2CID 37683420.
- ^ Jacoby, D. M. P.; Siriwat, P.; Freeman, R.; Carbone, C. (2015). «Is the scaling of swim speed in sharks driven by metabolism?». Biology Letters. 12 (10): 20150781. doi:10.1098/rsbl.2015.0781. PMC 4707698. PMID 26631246.
- ^ a b c Ferrón, H. G. (2017). «Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks». PLOS ONE. 12 (9): e0185185. Bibcode:2017PLoSO..1285185F. doi:10.1371/journal.pone.0185185. PMC 5609766. PMID 28938002.
- ^ Shimada, K.; Becker, M. A.; Griffiths, M. L. (2020). «Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to ‘off-the-scale’ gigantism of the megatooth shark, Otodus megalodon«. Historical Biology. 33 (11): 1–17. doi:10.1080/08912963.2020.1812598.
- ^ a b c d e f g Prothero, D. R. (2015). «Mega-Jaws». The Story of Life in 25 Fossils. New York, New York: Columbia University Press. pp. 96–110. ISBN 978-0-231-17190-8. OCLC 897505111.
- ^ Helfman, G.; Burgess, G. H. (2014). Sharks: The Animal Answer Guide. Baltimore, Maryland: Johns Hopkins University Press. p. 19. ISBN 978-1-4214-1310-5. OCLC 903293986.
- ^ Randall, John E. (1973). «Size of the Great White Shark (Carcharodon)». Science Magazine. 181 (4095): 169–170. Bibcode:1973Sci…181..169R. doi:10.1126/science.181.4095.169. PMID 17746627. S2CID 36607712.
- ^ Schembri, Patrick (1994). «Malta’s Natural Heritage». Natural Heritage. In: 105–124.
- ^ Papson, Stephen (1992). «Copyright: Cross the Fin Line of Terror». Journal of American Culture. 15 (4): 67–81. doi:10.1111/j.1542-734X.1992.1504_67.x.
- ^ Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ Shimada, Kenshu (2002). «The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)». Journal of Fossil Research. 35 (2): 28–33.
- ^ a b Pimiento, Catalina; Gerardo González-Barba; Dana J. Ehret; Austin J. W. Hendy; Bruce J. MacFadden; Carlos Jaramillo (2013). «Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama» (PDF). Journal of Paleontology. 87 (5): 755–774. doi:10.1666/12-117. S2CID 45662900. Archived from the original (PDF) on 29 October 2013.
- ^ Crane, B. (2017). «A Prehistoric Killer, Buried in Muck». The New Yorker. Retrieved 10 December 2017.
- ^ Mustain, A. (2011). «For Sale: World’s Largest Shark Jaws». LiveScience. Retrieved 31 August 2017.
- ^ a b Almgreen, S. E. Bendix (15 November 1983). «Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn». Bulletin of the Geological Society of Denmark. 32: 1–32. CiteSeerX 10.1.1.514.1782. doi:10.37570/bgsd-1983-32-01. S2CID 53311833. NAID 10012345550.
- ^ Reolid, M.; Molina, J. M. (2015). «Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain)». Estudios Geológicos. 71 (2): e032. doi:10.3989/egeol.41828.342.
- ^ Uyeno, T.; Sakamoto, O.; Sekine, H. (1989). «The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan». Saitama Museum of Natural History Bulletin. 7: 73–85.
- ^ Anderson, P.S.L.; Westneat, M. (2009). «A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)». Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1. S2CID 86203770.
- ^ Ballell, A.; Ferrón, H.G. (2021). «Biomechanical insights into the dentition of megatooth sharks (Lamniformes: Otodontidae)». Scientific Reports. 11 (1232): 1232. doi:10.1038/s41598-020-80323-z. PMC 7806677. PMID 33441828.
- ^ a b Stringer, G. L.; King, L. (2012). «Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana». New Mexico Museum of Natural History and Science, Bulletin. Vertebrate Corpolites. 57: 301.
- ^ «Megalodon Shark Facts and Information: The Details». fossilguy.com. Retrieved 18 September 2017.
- ^ Fitzgerald, Erich (2004). «A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia». Memoirs of Museum Victoria. 61 (2): 183–208. doi:10.24199/j.mmv.2004.61.12.
- ^ Löffler, N.; Fiebig, J.; Mulch, A.; Tütken, T.; Schmidt, B.C.; Bajnai, D.; Conrad, A.C.; Wacker, U.; Böttcher, M.E. (2019). «Refining the temperature dependence of the oxygen and clumped isotopic compositions of structurally bound carbonate in apatite». Geochimica et Cosmochimica Acta. 253: 19–38. Bibcode:2019GeCoA.253…19L. doi:10.1016/j.gca.2019.03.002. S2CID 107992832.
- ^ a b c Aguilera O.; Augilera E. R. D. (2004). «Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage». Caribbean Journal of Science. 40 (3): 362–368.
- ^ a b «Carcharocles megalodon«. Fossilworks. Retrieved 28 August 2017 from the Paleobiology Database.
{{cite web}}: CS1 maint: postscript (link) - ^ «Young Prince George gifted 23m-year-old tooth from extinct shark found in Malta». Times of Malta. 27 September 2020. Retrieved 28 September 2020.
- ^ Yun, C. (2020). «New example of Cosmopolitodus hastalis (Lamniformes, Lamnidae) from the Miocene South Korea». Zoodiversity. 54 (5): 433–438. doi:10.15407/zoo2020.05.433. S2CID 229274996.
- ^ Choi, Seung; Lee, Yuong-Nam (2017). «A review of vertebrate body fossils from the Korean Peninsula and perspectives». Geosciences Journal. 21 (6): 867–889. Bibcode:2017GescJ..21..867C. doi:10.1007/s12303-017-0040-6. ISSN 1226-4806. S2CID 133835817.
- ^ Martin, J. E.; Tacail, T.; Sylvain, A.; Catherine, G.; Vincent, B. (2015). «Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs». Chemical Geology. 415: 118–125. Bibcode:2015ChGeo.415..118M. doi:10.1016/j.chemgeo.2015.09.011.
- ^ a b c Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ a b Morgan, Gary S. (1994). «Whither the giant white shark?» (PDF). Paleontology Topics. 2 (3): 1–2. Archived from the original (PDF) on 22 July 2016.
- ^ Augilera, Orangel A.; García, Luis; Cozzuol, Mario A. (2008). «Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation». Paläontologische Zeitschrift. 82 (2): 204–208. doi:10.1007/BF02988410. ISSN 0038-2353. S2CID 84251638.
- ^ Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). «The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)». Journal of South American Earth Sciences. 73: 168–190. Bibcode:2017JSAES..73..168L. doi:10.1016/j.jsames.2016.12.010.
- ^ a b c d e f g h Lambert, O.; Bianucci, G.; Post, P.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. (2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020. S2CID 4369352.
- ^ Compagno, Leonard J. V. (1989). «Alternative life-history styles of cartilaginous fishes in time and space». Environmental Biology of Fishes. 28 (1–4): 33–75. doi:10.1007/BF00751027. S2CID 22527888.
- ^ Ferretti, Francesco; Boris Worm; Gregory L. Britten; Michael R. Heithaus; Heike K. Lotze1 (2010). «Patterns and ecosystem consequences of shark declines in the ocean» (PDF). Ecology Letters. 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. Archived from the original (PDF) on 6 July 2011. Retrieved 19 February 2011.
- ^ Gilbert, K.N.; Ivany, L.C.; Uhen, M.D. (2018). «Living fast and dying young: life history and ecology of a Neogene sperm whale». Journal of Vertebrate Paleontology. 38 (2): e1439038. doi:10.1080/02724634.2018.1439038. S2CID 89750852.
- ^ a b c d e Heyning, John; Dahlheim, Marilyn (1988). «Orcinus orca» (PDF). Mammalian Species (304): 1–9. doi:10.2307/3504225. JSTOR 3504225. S2CID 253914153. Archived from the original (PDF) on 5 December 2010.
- ^ Bianucci, Giovanni; Walter, Landini (2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society. 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x.
- ^ Lindberg, D. R.; Pyenson, N. D. (2006). «Evolutionary Patterns in Cetacea: Fishing Up Prey Size through Deep Time». Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 77. ISBN 978-0-520-24884-7.
- ^ Boessenecker, R. W. (2013). «A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans». Geodiversitas. 35 (4): 815–940. doi:10.5252/g2013n4a5. S2CID 85940452.
- ^ Bianucci, G. (1997). «Hemisyntrachelus cortesii (Cetacea, Delphinidae) from the Pliocene Sediments of Campore Quarry (Salsomaggiori Terme, Italy». Bollettino della Societa Paleontologica Italiana. 36 (1): 75–83).
- ^ Antunes, M.T.; Legoinha, P.; Balbing, A. (2015). «Megalodon, mako shark and planktonic foraminifera from the continental shelf off Portugal and their age». Geologica Acta. 13: 181–190.
- ^ «Paleoecology of Megalodon and the White Shark». Biology of Sharks and Rays. Retrieved 1 October 2017.
- ^ Tanke, Darren; Currie, Philip (1998). «Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence» (PDF). Gaia (15): 167–184.
- ^ a b Godfrey, S. J.; Altman, J. (2005). «A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon» (PDF). Jeffersoniana (16): 1–12.
- ^ STEPHEN J. GODFREY; JOHN R. NANCE; NORMAN L. RIKER (2021). «Otodus-bitten sperm whale tooth from the Neogene of the Coastal Eastern United States» (PDF). Acta Palaeontologica Polonica. 66 (3): 599–603.
- ^ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). «The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes». Journal of Mammalian Evolution. 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3. S2CID 90231.
- ^ Ehret D.J. (2010). «CHAPTER 5 – MACROEVOLUTION, AGE, AND GROWTH DETERMINATION OF THE MEGATOOTHED SHARKS (LAMNIFORMES: OTODONTIDAE)». Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes) (PDF). pp. 100–136.
- ^ S., Kenshu; Bonnan, M. F.; Becker, M. A.; Griffiths, M. L. (2021). «Ontogenetic growth pattern of the extinct megatooth shark Otodus megalodon—implications for its reproductive biology, development, and life expectancy». Historical Biology. 33 (12): 3254–3259. doi:10.1080/08912963.2020.1861608.
- ^ «Identifican en Canarias fósiles de ‘megalodón’, el tiburón más grande que ha existido» [Identifying Canary fossils of ‘megalodon’, the largest shark that ever lived] (in Spanish). Europa Press Noticias SA. 2013. Retrieved 29 August 2017.
- ^ Dulvy, N. K.; Reynolds, J. D. (1997). «Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays». Proceedings of the Royal Society B: Biological Sciences. 264 (1386): 1309–1315. Bibcode:1997RSPSB.264.1309D. doi:10.1098/rspb.1997.0181. PMC 1688595.
- ^ Godfrey, Stephen (2004). «The Ecphora» (PDF). The Newsletter of Calvert Marine Museum Fossil Club. 19 (1): 1–13. Archived from the original (PDF) on 10 December 2010.
- ^ Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (27 August 2010). «Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event». International Journal of Osteoarchaeology. 22 (3): 253–260. doi:10.1002/oa.1199.
- ^ a b Pimiento, C.; Griffin, J. N.; Clements, C. F.; Silvestro, D.; Varela, S.; Uhen, M. D.; Jaramillo, C. (2017). «The Pleistocene Marine Megafauna Extinction and its Impact on Functional Diversity». Nature Ecology and Evolution. 1 (8): 1100–1106. doi:10.1038/s41559-017-0223-6. PMID 29046566. S2CID 3639394.
- ^ a b c d «The Extinction of Megalodon». Biology of Sharks and Rays. Retrieved 31 August 2017.
- ^ Reilly, Michael (29 September 2009). «Prehistoric Shark Nursery Spawned Giants». Discovery News. Archived from the original on 10 March 2012. Retrieved 23 November 2013.
- ^ a b c Allmon, Warren D.; Steven D. Emslie; Douglas S. Jones; Gary S. Morgan (2006). «Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms». The Journal of Geology. 104 (2): 143–162. Bibcode:1996JG….104..143A. doi:10.1086/629811. S2CID 128418299.
- ^ a b Collareta, A.; Lambert, O.; Landini, W.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ Dooly A.C.; Nicholas C. F.; Luo Z. X. (2006). «The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)». Journal of Vertebrate Paleontology. 24 (2): 453–463. doi:10.1671/2401. JSTOR 4524731. S2CID 84970052.
- ^ a b c McCormack, Jeremy; Griffiths, Michael L.; Kim, Sora L.; Shimada, Kenshu; Karnes, Molly; Maisch, Harry; Pederzani, Sarah; Bourgon, Nicolas; Jaouen, Klervia; Becker, Martin A.; Jöns, Niels (31 May 2022). «Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes». Nature Communications. 13 (1): 2980. Bibcode:2022NatCo..13.2980M. doi:10.1038/s41467-022-30528-9. ISSN 2041-1723. PMC 9156768. PMID 35641494. S2CID 249235478.
- ^ a b c d Citron, Sara; Geisler, Jonathan H.; Alberto, Collareta; Giovanni, Bianucci (2022). «Systematics, phylogeny and feeding behavior of the oldest killer whale: a reappraisal of Orcinus citoniensis (Capellini, 1883) from the Pliocene of Tuscany (Italy)». Bollettino della Società Paleontologica Italiana. 61 (2): 167–186. doi:10.4435/BSPI.2022.13.
- ^ «Huge Tooth Reveals Prehistoric Moby Dick in Melbourne». Australasian Science Magazine. Retrieved 24 April 2016.
- ^ «Move over Moby Dick: Meet Melbourne’s own mega whale». The Sydney Morning Herald. 21 April 2016.
- ^ Govender, R (2021). «Early Pliocene fossil cetaceans from Hondeklip Bay, Namaqualand, South Africa». Historical Biology. 33 (4): 574–593. doi:10.1080/08912963.2019.1650273. S2CID 202019648.
- ^ Hampe, O. (2006). «Middle/late Miocene hoplocetine sperm whale remains (Odontoceti: Physeteridae) of North Germany with an emended classification of the Hoplocetinae». Fossil Record. 9 (1): 61–86. doi:10.1002/mmng.200600002.
- ^ a b Antunes, Miguel Telles; Balbino, Ausenda Cáceres (2010). «The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic». Revista Española de Paleontología. 25 (1): 1–6.
- ^ Kast, Emma R.; Griffiths, Michael L.; Kim, Sora. L.; Rao, Zixuan C.; Shimada, Kensu; Becker, Martin A.; Maisch, Harry M.; Eagle, Robert A.; Clarke, Chelesia A.; Neumann, Allison N.; Karnes, Molly E.; Lüdecke, Tina; Leichliter, Jennifer N.; Martínez-García, Alfredo; Akhtar, Alliya A.; Wang, Xingchen T.; Haug, Gerald H.; Sigman, Daniel M. (22 June 2022). «Cenozoic megatooth sharks occupied extremely high trophic positions». Science Advances. 8 (25): eabl6529. Bibcode:2022SciA….8L6529K. doi:10.1126/sciadv.abl6529. PMC 9217088. PMID 35731884.
- ^ Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (2017). «Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics». Proceedings of the Royal Society B: Biological Sciences. 284 (1855): 20170546. doi:10.1098/rspb.2017.0546. PMC 5454272. PMID 28539520.
- ^ Sylvain, Adnet; A. C. Balbino; M. T. Antunes; J. M. Marín-Ferrer (2010). «New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean». Neues Jahrbuch für Geologie und Paläontologie. 256 (1): 7–16. doi:10.1127/0077-7749/2009/0029.
- ^ Weinstock, J. A. (2014). The Ashgate Encyclopedia of Literary and Cinematic Monsters. Farnham, United Kingdom: Routledge. pp. 107–108. ISBN 978-1-4094-2562-5. OCLC 874390267.
- ^ a b Guimont, Edward (5 October 2021). «The Megalodon: A Monster of the New Mythology». M/C Journal. 24 (5). doi:10.5204/mcj.2793. ISSN 1441-2616. S2CID 241813307.
- ^ Roesch, B. S. (1998). «A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles megalodon«. The Cryptozoology Review. 3 (2): 14–24.
- ^ «Does Megalodon Still Live?». Biology of Sharks and Rays. Retrieved 2 October 2017.
- ^ «The Meg (2018)». Box Office Mojo. Retrieved 22 December 2017.
- ^ Sid Bennett (director) (27 May 2012). Mermaids: The Body Found (Motion picture). Animal Planet.
- ^ «Shark Week ‘Megalodon: The Monster Shark Lives’ Tries To Prove Existence Of Prehistoric Shark (VIDEO)». Huff Post Green. 5 August 2013. Retrieved 11 August 2013.
- ^ Winston, B.; Vanstone, G.; Chi, W. (2017). «A Walk in the Woods». The Act of Documenting: Documentary Film in the 21st Century. New York, New York: Bloomsbury Publishing. ISBN 978-1-5013-0918-2. OCLC 961183719.
- ^ Flanagin, J. (2014). «Sorry, Fans. Discovery Has Jumped the Shark Week». New York Times. Retrieved 16 August 2014.
- ^ Shiffman, David (15 August 2014). «Shark Week Is Lying Again About Monster Megalodon Sharks». Slate Magazine. Retrieved 31 July 2022.
- ^ O’Connell, Mikey (5 August 2013). «TV Ratings: Shark Week Hits Record Highs With Fake ‘Megalodon’ Doc». The Hollywood Reporter. Retrieved 31 July 2022.
- ^ «Fossil, Fossilized Teeth of the Megalodon Shark | NCpedia». ncpedia.org. Retrieved 17 October 2019.
Further reading
- Dickson, K. A.; Graham, J. B. (November–December 2004). «Evolution and consequences of endothermy in fishes». Physiological and Biochemical Zoology. 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772. S2CID 40104003.
- Kent, Bretton W. (1994). Fossil Sharks of the Chesapeake Bay Region. Columbia, Md.: Egan Rees & Boyer. ISBN 978-1-881620-01-3. OCLC 918266672.
External links
- The rise of super predatory sharks
- Carcharocles: Extinct Megatoothed shark
- Paleontological videos
- Paleontologist Mark Renz shows one of the largest megalodon teeth discovered on YouTube
- Shark Week Special on megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about megalodon on YouTube (featuring expert Dana Ehret)
- Lamniform sharks: 110 million years of ocean supremacy on YouTube (featuring expert Mikael Siverson)
- The Rise and Fall of the Neogene Giant Sharks on YouTube (featuring expert Bretton Kent)
- Moore, Kallie (19 December 2018). «Why Megalodon (Definitely) Went Extinct». PBS Eons. Archived from the original on 14 November 2021 – via YouTube.
| В Википедии есть статья «мегалодон». |
Содержание
- 1 Русский
- 1.1 Морфологические и синтаксические свойства
- 1.2 Произношение
- 1.3 Семантические свойства
- 1.3.1 Значение
- 1.3.2 Синонимы
- 1.3.3 Антонимы
- 1.3.4 Гиперонимы
- 1.3.5 Гипонимы
- 1.4 Родственные слова
- 1.5 Этимология
- 1.6 Фразеологизмы и устойчивые сочетания
- 1.7 Перевод
- 1.8 Библиография
Русский[править]
| В Викиданных есть лексема мегалодон (L126178). |
Морфологические и синтаксические свойства[править]
| падеж | ед. ч. | мн. ч. |
|---|---|---|
| Им. | мегалодо́н | мегалодо́ны |
| Р. | мегалодо́на | мегалодо́нов |
| Д. | мегалодо́ну | мегалодо́нам |
| В. | мегалодо́на | мегалодо́нов |
| Тв. | мегалодо́ном | мегалодо́нами |
| Пр. | мегалодо́не | мегалодо́нах |
ме—га—ло—до́н
Существительное, одушевлённое, мужской род, 2-е склонение (тип склонения 1a по классификации А. А. Зализняка).
Корень: —.
Произношение[править]
- МФА: ед. ч. [mʲɪɡəɫɐˈdon], мн. ч. [mʲɪɡəɫɐˈdonɨ]
Семантические свойства[править]
Значение[править]
- ихтиол., палеонт. вымершая акула из семейства Otodontidae, существовавшая в миоцене и плиоцене, одна из самых больших рыб всех времён ◆ Отсутствует пример употребления (см. рекомендации).
Синонимы[править]
- —
Антонимы[править]
- —
Гиперонимы[править]
- акула, рыба
Гипонимы[править]
- —
Родственные слова[править]
| Ближайшее родство | |
Этимология[править]
От ??
Фразеологизмы и устойчивые сочетания[править]
Перевод[править]
| Список переводов | |
Библиография[править]
|
|
Для улучшения этой статьи желательно:
|
| Megalodon
Temporal range: Early Miocene–Early Pliocene, c. 23–3.6 Ma PreꞒ Ꞓ O S D C P T J K Pg N |
|
|---|---|
|
|
|
| Model of megalodon jaws at the American Museum of Natural History | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Otodontidae |
| Genus: | †Otodus |
| Species: |
†O. megalodon |
| Binomial name | |
| †Otodus megalodon
(Agassiz, 1843)[1] |
|
| Synonyms[2][3][4][5][6] | |
|
List of synonyms
|
Megalodon (Otodus megalodon),[6][7][8] meaning «big tooth», is an extinct species of mackerel shark that lived approximately 23 to 3.6 million years ago (Mya), from the Early Miocene to the Pliocene epochs.[9] It was formerly thought to be a member of the family Lamnidae and a close relative of the great white shark (Carcharodon carcharias). However, it is now classified into the extinct family Otodontidae, which diverged from the great white shark during the Early Cretaceous.
While regarded as one of the largest and most powerful predators to have ever lived, the megalodon is only known from fragmentary remains, and its appearance and maximum size are uncertain. Scientists differ on whether it would have more closely resembled a stockier version of the great white shark, the whale shark (Rhincodon typus), the basking shark (Cetorhinus maximus) or the sand tiger shark (Carcharias taurus). The most recent estimate with the least error range suggests a maximum length estimate up to 20.3 meters (67 ft),[10] although the modal lengths are estimated at 10.5 meters (34 ft).[11][12] Extrapolation from a vertebral centra with dimensions based on the great white shark suggests that a megalodon about 16 meters (52 ft) long weighs up to 48 metric tons (53 short tons), 17 meters (56 ft) long weighs up to 59 metric tons (65 short tons), and 20.3 meters (67 ft) long (the maximum length) weighs up to 103 metric tons (114 short tons).[13][14] Extrapolating from a vertebral column and reconstructing a 3D model with dimensions based on all extant lamnid sharks suggests that a 16-meter-long (52 ft) individual may have been much larger than previous estimates, reaching an excess of 61.5 metric tons (67.8 short tons) in body mass; an individual of this size would have needed to consume 98,175 kcal per day.[15] Their teeth were thick and robust, built for grabbing prey and breaking bone, and their large jaws could exert a bite force of up to 108,500 to 182,200 newtons (24,400 to 41,000 lbf).[14]
Megalodon probably had a major impact on the structure of marine communities. The fossil record indicates that it had a cosmopolitan distribution. It probably targeted large prey, such as whales, seals and sea turtles. Juveniles inhabited warm coastal waters and fed on fish and small whales. Unlike the great white, which attacks prey from the soft underside, megalodon probably used its strong jaws to break through the chest cavity and puncture the heart and lungs of its prey.
The animal faced competition from whale-eating cetaceans, such as Livyatan and other macroraptorial sperm whales and possibly smaller ancestral killer whales. As the shark preferred warmer waters, it is thought that oceanic cooling associated with the onset of the ice ages, coupled with the lowering of sea levels and resulting loss of suitable nursery areas, may have also contributed to its decline. A reduction in the diversity of baleen whales and a shift in their distribution toward polar regions may have reduced megalodon’s primary food source. The shark’s extinction coincides with a gigantism trend in baleen whales.
Taxonomy
Naming
The depiction of a shark’s head by Nicolas Steno in his work The Head of a Shark Dissected
According to Renaissance accounts, gigantic triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolas Steno, who recognized them as shark teeth, and famously produced a depiction of a shark’s head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a megalodon tooth.[16][17][18]
Swiss naturalist Louis Agassiz gave this shark its initial scientific name, Carcharodon megalodon, in his 1843 work Recherches sur les poissons fossiles, based on tooth remains.[1][19] English paleontologist Edward Charlesworth in his 1837 paper used the name Carcharias megalodon, while citing Agassiz as the author, indicating that Agassiz described the species prior to 1843. English paleontologist Charles Davies Sherborn in 1928 listed an 1835 series of articles by Agassiz as the first scientific description of the shark.[20] The specific name megalodon translates to «big tooth», from Ancient Greek: μέγας, romanized: (mégas), lit. ‘big, mighty’ and ὀδούς (odoús), «tooth».[21][22] The teeth of megalodon are morphologically similar to those of the great white shark (Carcharodon carcharias), and on the basis of this observation, Agassiz assigned megalodon to the genus Carcharodon.[19]
There was one apparent description of the shark in 1881 classifying it as Selache manzonii.[23]
Evolution
While the earliest megalodon remains have been reported from the Late Oligocene, around 28 million years ago (Mya),[24][25] there is disagreement as to when it appeared, with dates ranging to as young as 16 mya.[26] It has been thought that megalodon became extinct around the end of the Pliocene, about 2.6 Mya;[26][27] claims of Pleistocene megalodon teeth, younger than 2.6 million years old, are considered unreliable.[27] A 2019 assessment moves the extinction date back to earlier in the Pliocene, 3.6 Mya.[28]
Megalodon is now considered to be a member of the family Otodontidae, genus Otodus, as opposed to its previous classification into Lamnidae, genus Carcharodon.[26][12][27][6][7] Megalodon’s classification into Carcharodon was due to dental similarity with the great white shark, but most authors currently believe that this is due to convergent evolution. In this model, the great white shark is more closely related to the extinct broad-toothed mako (Isurus hastalis) than to megalodon, as evidenced by more similar dentition in those two sharks; megalodon teeth have much finer serrations than great white shark teeth. The great white shark is more closely related to the mako shark (Isurus spp.), with a common ancestor around 4 Mya.[19][29] Proponents of the former model, wherein megalodon and the great white shark are more closely related, argue that the differences between their dentition are minute and obscure.[30]: 23–25
The genus Carcharocles currently contains four species: C. auriculatus, C. angustidens, C. chubutensis, and C. megalodon.[31]: 30–31 The evolution of this lineage is characterized by the increase of serrations, the widening of the crown, the development of a more triangular shape, and the disappearance of the lateral cusps.[31]: 28–31 [32] The evolution in tooth morphology reflects a shift in predation tactics from a tearing-grasping bite to a cutting bite, likely reflecting a shift in prey choice from fish to cetaceans.[33] Lateral cusplets were finally lost in a gradual process that took roughly 12 million years during the transition between C. chubutensis and C. megalodon.[33] The genus was proposed by D. S. Jordan and H. Hannibal in 1923 to contain C. auriculatus. In the 1980s, megalodon was assigned to Carcharocles.[19][31]: 30 Before this, in 1960, the genus Procarcharodon was erected by French ichthyologist Edgard Casier, which included those four sharks and was considered separate from the great white shark. It is now considered a junior synonym of Carcharocles.[31]: 30 The genus Palaeocarcharodon was erected alongside Procarcharodon to represent the beginning of the lineage, and, in the model wherein megalodon and the great white shark are closely related, their last common ancestor. It is believed to be an evolutionary dead-end and unrelated to the Carcharocles sharks by authors who reject that model.[30]: 70
The great white shark (Carcharodon carcharias) and megalodon were previously thought to be close relatives.[19][29]
Another model of the evolution of this genus, also proposed by Casier in 1960, is that the direct ancestor of the Carcharocles is the shark Otodus obliquus, which lived from the Paleocene through the Miocene epochs, 60 to 13 Mya.[29][32] The genus Otodus is ultimately derived from Cretolamna, a shark from the Cretaceous period.[6][34] In this model, O. obliquus evolved into O. aksuaticus, which evolved into C. auriculatus, and then into C. angustidens, and then into C. chubutensis, and then finally into C. megalodon.
Another model of the evolution of Carcharocles, proposed in 2001 by paleontologist Michael Benton, is that the three other species are actually a single species of shark that gradually changed over time between the Paleocene and the Pliocene, making it a chronospecies.[31]: 17 [25][35] Some authors suggest that C. auriculatus, C. angustidens, and C. chubutensis should be classified as a single species in the genus Otodus, leaving C. megalodon the sole member of Carcharocles.[25][36]
The genus Carcharocles may be invalid, and the shark may actually belong in the genus Otodus, making it Otodus megalodon.[4] A 1974 study on Paleogene sharks by Henri Cappetta erected the subgenus Megaselachus, classifying the shark as Otodus (Megaselachus) megalodon, along with O. (M.) chubutensis. A 2006 review of Chondrichthyes elevated Megaselachus to genus, and classified the sharks as Megaselachus megalodon and M. chubutensis.[4] The discovery of fossils assigned to the genus Megalolamna in 2016 led to a re-evaluation of Otodus, which concluded that it is paraphyletic, that is, it consists of a last common ancestor but it does not include all of its descendants. The inclusion of the Carcharocles sharks in Otodus would make it monophyletic, with the sister clade being Megalolamna.[6]
The cladogram below represents the hypothetical relationships between megalodon and other sharks, including the great white shark. Modified from Shimada et al. (2016),[6] Ehret et al., (2009),[29] and the findings of Siversson et al. (2013).[38][39]
Biology
Appearance
One interpretation on how megalodon appeared was that it was a robust-looking shark, and may have had a similar build to the great white shark. The jaws may have been blunter and wider than the great white, and the fins would have also been similar in shape, though thicker due to its size. It may have had a pig-eyed appearance, in that it had small, deep-set eyes.[40]
Another interpretation is that megalodon bore a similarity to the whale shark (Rhincodon typus) or the basking shark (Cetorhinus maximus). The tail fin would have been crescent-shaped, the anal fin and second dorsal fin would have been small, and there would have been a caudal keel present on either side of the tail fin (on the caudal peduncle). This build is common in other large aquatic animals, such as whales, tuna, and other sharks, in order to reduce drag while swimming. The head shape can vary between species as most of the drag-reducing adaptations are toward the tail-end of the animal.[31]: 35–36
Sculpture in the Museum of Evolution in Puebla, Mexico
Since Carcharocles is derived from Otodus, and the two had teeth that bear a close similarity to those of the sand tiger shark (Carcharias taurus), megalodon may have had a build more similar to the sand tiger shark than to other sharks. This is unlikely since the sand tiger shark is a carangiform swimmer which requires faster movement of the tail for propulsion through the water than the great white shark, a thunniform swimmer.[31]: 35–36 [41]
Size
Due to fragmentary remains, there have been many contradictory size estimates for megalodon, as they can only be drawn from fossil teeth and vertebrae.[42]: 87 [43] The great white shark has been the basis of reconstruction and size estimation, as it is regarded as the best analogue to megalodon. Several total length estimation methods have been produced from comparing megalodon teeth and vertebrae to those of the great white.[40][44][10][7]
Size comparison of the great white and whale shark to estimates for megalodon
Proportions of megalodon at lengths of 3 m (10 ft), 8 m (26 ft), and 16 m (52 ft), extrapolated from extant relatives, with a 1.65 m (5 ft 5 in) diver
Megalodon size estimates vary depending on the method used, with maximum total length estimates ranging from 14.2–20.3 meters (47–67 ft).[40][10][7] A 2015 study estimated the average total body length at 10.5 meters (34 ft), calculated from 544 megalodon teeth, found throughout geological time and geography, including adults and juveniles.[11][12] In comparison, large great white sharks are generally around 6 meters (20 ft) in length, with a few contentious reports suggesting larger sizes.[45][46][40] The whale shark is the largest living fish, with one large female reported with a precaudal length of 15 meters (49 ft) and an estimated total length of 18.8 meters (62 ft).[45][47] It is possible that different populations of megalodon around the globe had different body sizes and behaviors due to different ecological pressures.[12] Megalodon is thought to have been the largest macropredatory shark that ever lived.[40]
«A C. megalodon about 16 meters long would have weighed about 48 metric tons (53 tons). A 17-meter (56-foot) C. megalodon would have weighed about 59 metric tons (65 tons), and a 20.3-meter (67 foot) monster would have topped off at 103 metric tons (114 tons).»[13]
In his 2015 book, The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution, Donald Prothero proposed the body mass estimates for different individuals of different length by extrapolating from a vertebral centra based on the dimensions of the great white,[13] a methodology also used for the 2008 study which supports the maximum mass estimate.[14]
In 2020, Cooper and his colleagues reconstructed a 2D model of megalodon based on the dimensions of all the extant lamnid sharks and suggested that a 16 meters (52 ft) long megalodon would have had a 4.65 m (15.3 ft) long head, 1.41 m (4 ft 8 in) tall gill slits, a 1.62 m (5 ft 4 in) tall dorsal fin, 3.08 m (10 ft 1 in) long pectoral fins, and a 3.85 m (12 ft 8 in) tall tail fin.[8] In 2022, Cooper and his colleagues also reconstructed a 3D model with the same basis as the 2020 study, resulting in a body mass estimate of 61.56 metric tons (67.86 short tons) for a 16 meters (52 ft) long megalodon (higher than the previous estimates); a vertebral column specimen named IRSNB P 9893 (formerly IRSNB 3121), belonging to a 46 year old individual from Belgium, was used for extrapolation. An individual of this size would have required 98,175 kcal per day, 20 times more than what the adult great white requires.[15]
Mature male megalodon may have had a body mass of 12.6 to 33.9 metric tons (13.9 to 37.4 short tons), and mature females may have been 27.4 to 59.4 metric tons (30.2 to 65.5 short tons), assuming that males could range in length from 10.5 to 14.3 meters (34 to 47 ft) and females 13.3 to 17 meters (44 to 56 ft).[40]
A 2015 study linking shark size and typical swimming speed estimated that megalodon would have typically swum at 18 kilometers per hour (11 mph)–assuming that its body mass was typically 48 metric tons (53 short tons)–which is consistent with other aquatic creatures of its size, such as the fin whale (Balaenoptera physalus) which typically cruises at speeds of 14.5 to 21.5 km/h (9.0 to 13.4 mph).[48] In 2022, Cooper and his colleagues converted this calculation into relative cruising speed (body lengths per second), resulting in an mean absolute cruising speed of 5 kilometers per hour (3.1 mph) and a mean relative cruising speed of 0.09 body lengths per second for a 16 meters (52 ft) long megalodon; the authors found their mean absolute cruising speed to be faster than any extant lamnid sharks and their mean relative cruising speed to be slower, consistent with previous estiamtes.[15]
Its large size may have been due to climatic factors and the abundance of large prey items, and it may have also been influenced by the evolution of regional endothermy (mesothermy) which would have increased its metabolic rate and swimming speed. The otodontid sharks have been considered to have been ectotherms, so on that basis megalodon would have been ectothermic. However, the largest contemporary ectothermic sharks, such as the whale shark, are filter feeders, while lamnids are now known to be regional endotherms, implying some metabolic correlations with a predatory lifestyle. These considerations, as well as tooth oxygen isotopic data and the need for higher burst swimming speeds in macropredators of endothermic prey than ectothermy would allow, imply that otodontids, including megalodon, were probably regional endotherms.[49]
In 2020, Shimada and colleagues suggested large size was instead due to intrauterine cannibalism, where the larger fetus eats the smaller fetus, resulting in progressively larger and larger fetuses, requiring the mother to attain even greater size as well as caloric requirements which would have promoted endothermy. Males would have needed to keep up with female size in order to still effectively copulate (which probably involved latching onto the female with claspers, like modern cartilaginous fish).[50]
Maximum estimates
The first attempt to reconstruct the jaw of megalodon was made by Bashford Dean in 1909, displayed at the American Museum of Natural History. From the dimensions of this jaw reconstruction, it was hypothesized that megalodon could have approached 30 meters (98 ft) in length. Dean had overestimated the size of the cartilage on both jaws, causing it to be too tall.[51][52]
Tooth compared to hand
In 1973, John E. Randall, an ichthyologist, used the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) to measure the length of the shark, yielding a maximum length of about 13 meters (43 ft).[53] However, tooth enamel height does not necessarily increase in proportion to the animal’s total length.[30]: 99
In 1994, marine biologists Patrick J. Schembri and Stephen Papson opined that O. megalodon may have approached a maximum of around 24 to 25 meters (79 to 82 ft) in total length.[54][55]
In 1996, shark researchers Michael D. Gottfried, Leonard Compagno, and S. Curtis Bowman proposed a linear relationship between the great white shark’s total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in meters = − (0.096) × [UA maximum height (mm)]-(0.22).[56][40] Using this tooth height regression equation, the authors estimated a total length of 15.9 meters (52 ft) based on a tooth 16.8 centimeters (6.6 in) tall, which the authors considered a conservative maximum estimate. They also compared the ratio between the tooth height and total length of large female great whites to the largest megalodon tooth. A 6-meter (20 ft) long female great white, which the authors considered the largest ‘reasonably trustworthy’ total length, produced an estimate of 16.8 meters (55 ft). However, based on the largest female great white reported, at 7.1 meters (23 ft), they estimated a maximum estimate of 20.2 meters (66 ft).[40]
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimeter (0.39 in) of root width, there are approximately 1.4 meters (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah’s possession had a root width of about 12 centimeters (4.7 in), which yielded 16.5 meters (54 ft) in total length.[31]: 88
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length after conducting anatomical analysis of several specimens, allowing any sized tooth to be used. Shimada stated that the previously proposed methods were based on a less-reliable evaluation of the dental homology between megalodon and the great white shark, and that the growth rate between the crown and root is not isometric, which he considered in his model. Using this model, the upper anterior tooth possessed by Gottfried and colleagues corresponded to a total length of 15 meters (49 ft).[57] Among several specimens found in the Gatún Formation of Panama, one upper lateral tooth was used by other researchers to obtain a total length estimate of 17.9 meters (59 ft) using this method.[36][58]
In 2019, Shimada revisited the size of megalodon and discouraged using non-anterior teeth for estimations, noting that the exact position of isolated non-anterior teeth is difficult to identify. Shimada provided maximum total length estimates using the largest anterior teeth available in museums. The tooth with the tallest crown height known to Shimada, NSM PV-19896, produced a total length estimate of 14.2 meters (47 ft). The tooth with the tallest total height, FMNH PF 11306, was reported at 16.8 centimeters (6.6 in). However, Shimada remeasured the tooth and found it actually to measure 16.2 centimeters (6.4 in). Using the total height tooth regression equation proposed by Gottfried and colleagues produced an estimate of 15.3 meters (50 ft).[7][10]
In 2021, Victor J. Perez, Ronny M. Leder, and Teddy Badaut proposed a method of estimating total length of megalodon from the sum of the tooth crown widths. Using more complete megalodon dentitions, they reconstructed the dental formula and then made comparisons to living sharks. The researchers noted that the 2002 Shimada crown height equations produce wildly varying results for different teeth belonging to the same shark (range of error of ± 9 metres (30 ft)), casting doubt on some of the conclusions of previous studies using that method. Using the largest tooth available to the authors, GHC 6, with a crown width of 13.3 centimeters (5.2 in), they estimated a maximum body length of approximately 20 meters (66 ft), with a range of error of approximately ± 3.5 metres (11 ft).[10] This maximum length estimate was also supported by Cooper and his colleagues in 2022.[15]
There are anecdotal reports of teeth larger than those found in museum collections.[7] Gordon Hubbell from Gainesville, Florida, possesses an upper anterior megalodon tooth whose maximum height is 18.4 centimeters (7.25 in), one of the largest known tooth specimens from the shark.[59] In addition, a 2.7-by-3.4-meter (9 by 11 ft) megalodon jaw reconstruction developed by fossil hunter Vito Bertucci contains a tooth whose maximum height is reportedly over 18 centimeters (7 in).[60]
Teeth and bite force
The most common fossils of megalodon are its teeth. Diagnostic characteristics include a triangular shape, robust structure, large size, fine serrations, a lack of lateral denticles, and a visible V-shaped neck (where the root meets the crown).[30]: 55 [36] The tooth met the jaw at a steep angle, similar to the great white shark. The tooth was anchored by connective tissue fibers, and the roughness of the base may have added to mechanical strength.[61] The lingual side of the tooth, the part facing the tongue, was convex; and the labial side, the other side of the tooth, was slightly convex or flat. The anterior teeth were almost perpendicular to the jaw and symmetrical, whereas the posterior teeth were slanted and asymmetrical.[62]
Megalodon teeth can measure over 180 millimeters (7.1 in) in slant height (diagonal length) and are the largest of any known shark species,[31]: 33 implying it was the largest of all macropredatory sharks.[40] In 1989, a nearly complete set of megalodon teeth was discovered in Saitama, Japan. Another nearly complete associated megalodon dentition was excavated from the Yorktown Formations in the United States, and served as the basis of a jaw reconstruction of megalodon at the National Museum of Natural History (USNM). Based on these discoveries, an artificial dental formula was put together for megalodon in 1996.[30]: 55 [63]
The dental formula of megalodon is: 2.1.7.43.0.8.4. As evident from the formula, megalodon had four kinds of teeth in its jaws: anterior, intermediate, lateral, and posterior. Megalodon’s intermediate tooth technically appears to be an upper anterior and is termed as «A3» because it is fairly symmetrical and does not point mesially (side of the tooth toward the midline of the jaws where the left and right jaws meet). Megalodon had a very robust dentition,[30]: 20–21 and had over 250 teeth in its jaws, spanning 5 rows.[31]: iv It is possible that large megalodon individuals had jaws spanning roughly 2 meters (6.6 ft) across.[31]: 129 The teeth were also serrated, which would have improved efficiency in cutting through flesh or bone.[19][31]: 1 The shark may have been able to open its mouth to a 75° angle, though a reconstruction at the USNM approximates a 100° angle.[40]
In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5-meter (8.2 ft) long specimen, and then isometrically scaled the results for its maximum size and the conservative minimum and maximum body mass of megalodon. They placed the bite force of the latter between 108,514 to 182,201 newtons (24,395 to 40,960 lbf) in a posterior bite, compared to the 18,216 newtons (4,095 lbf) bite force for the largest confirmed great white shark, and 7,400 newtons (1,700 lbf) for the placoderm fish Dunkleosteus. In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the force generated, which would probably have caused the total force experienced by prey to be higher than the estimate.[14][64]
In 2021, Antonio Ballell and Humberto Ferrón used Finite Element Analysis modeling to examine the stress distribution of three types of megalodon teeth and closely related mega-toothed species when exposed to anterior and lateral forces, the latter of which would be generated when a shark shakes its head to tear through flesh. The resulting simulations identified higher levels of stress in megalodon teeth under lateral force loads compared to its precursor species such as O. obliquus and O. angusteidens when tooth size was removed as a factor. This suggests that megalodon teeth were of a different functional significance than previously expected, challenging prior interpretations that megalodon’s dental morphology was primarily driven by a dietary shift towards marine mammals. Instead, the authors proposed that it was a byproduct of an increase in body size caused by heterochronic selection.[65]
Internal anatomy
Megalodon is represented in the fossil record by teeth, vertebral centra, and coprolites.[40][66] As with all sharks, the skeleton of megalodon was formed of cartilage rather than bone; consequently most fossil specimens are poorly preserved.[67] To support its large dentition, the jaws of megalodon would have been more massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. Its chondrocranium, the cartilaginous skull, would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size.[40]
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp Basin, Belgium, in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimeters (2.2 in) to 155 millimeters (6 in) in diameter. The shark’s vertebrae may have gotten much bigger, and scrutiny of the specimen revealed that it had a higher vertebral count than specimens of any known shark, possibly over 200 centra; only the great white approached it.[40] Another partially preserved vertebral column of a megalodon was excavated from the Gram Formation in Denmark in 1983, which comprises 20 vertebral centra, with the centra ranging from 100 millimeters (4 in) to 230 millimeters (9 in) in diameter.[61]
The coprolite remains of megalodon are spiral-shaped, indicating that the shark may have had a spiral valve, a corkscrew-shaped portion of the lower intestines, similar to extant lamniform sharks. Miocene coprolite remains were discovered in Beaufort County, South Carolina, with one measuring 14 cm (5.5 in).[66]
Gottfried and colleagues reconstructed the entire skeleton of megalodon, which was later put on display at the Calvert Marine Museum in the United States and the Iziko South African Museum.[40][32] This reconstruction is 11.3 meters (37 ft) long and represents a mature male,[40]: 61 based on the ontogenetic changes a great white shark experiences over the course of its life.[40]: 65
Paleobiology
Range and habitat
Megalodon had a cosmopolitan distribution;[26][58] its fossils have been excavated from many parts of the world, including Europe, Africa, the Americas, and Australia.[30]: 67 [68] It most commonly occurred in subtropical to temperate latitudes.[26][30]: 78 It has been found at latitudes up to 55° N; its inferred tolerated temperature range was 1–24 °C (34–75 °F).[note 1] It arguably had the capacity to endure such low temperatures due to mesothermy, the physiological capability of large sharks to maintain a higher body temperature than the surrounding water by conserving metabolic heat.[26]
Megalodon inhabited a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult megalodon were not abundant in shallow water environments, and mostly inhabited offshore areas. Megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.[31]: 33 [70]
Fossil remains show a trend for specimens to be larger on average in the Southern Hemisphere than in the Northern, with mean lengths of 11.6 and 9.6 meters (38 and 31 ft), respectively; and also larger in the Pacific than the Atlantic, with mean lengths of 10.9 and 9.5 meters (36 and 31 ft) respectively. They do not suggest any trend of changing body size with absolute latitude, or of change in size over time (although the Carcharocles lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 meters (34 ft), with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.[12]
Locations of fossils
Megalodon had a global distribution and fossils of the shark have been found in many places around the world, bordering all oceans of the Neogene.[71]
| Epoch | Formation | State | Continent |
|---|---|---|---|
| Pliocene | Luanda Formation | Africa | |
| Africa | |||
| Africa | |||
| Castell’Arquato Formation | Europe | ||
| Arenas de Huelva Formation | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Touril Complex Formation | Europe | ||
| Red Crag Formation | Europe | ||
| San Mateo Formation | North America | ||
| Towsley Formation | North America | ||
| Bone Valley Formation | North America | ||
| Tamiami Formation | North America | ||
| Yorktown Formation | North America | ||
| Highlands Formation | North America | ||
| Refugio Formation | North America | ||
| San Diego Formation | North America | ||
| Tirabuzon Formation | North America | ||
| Onzole Formation | South America | ||
| Paraguaná Formation | South America | ||
| Black Rock Sandstone | Oceania | ||
| Cameron Inlet Formation | Oceania | ||
| Grange Burn Formation | Oceania | ||
| Loxton Sand Formation | Oceania | ||
| Whaler’s Bluff Formation | Oceania | ||
| Tangahoe Formation | Oceania | ||
| Miocene | |||
| Africa | |||
| Madagascar Basin | Africa | ||
| Africa | |||
| Varswater Formation | Africa | ||
| Baripada Limestone | Asia | ||
| Arakida Formation | Asia | ||
| Bihoku Group | Asia | ||
| Fujina Formation | Asia | ||
| Hannoura Formation | Asia | ||
| Hongo Formation | Asia | ||
| Horimatsu Formation | Asia | ||
| Ichishi Formation | Asia | ||
| Kurahara Formation | Asia | ||
| Maenami Formation | Asia | ||
| Matsuyama Group | Asia | ||
| Sekinobana Formation | Asia | ||
| Suso Formation | Asia | ||
| Takakubo Formation | Asia | ||
| Tonokita Formation | Asia | ||
| Tsurushi Formation | Asia | ||
| Wajimazaki Formation | Asia | ||
| Yoshii Formation | Asia | ||
| Asia | |||
| Burgeschleinitz Formation | Europe | ||
| Melker Sand Formation | Europe | ||
| Rzehakia Formation | Europe | ||
| Weissenegg Formation | Europe | ||
| Antwerpen Sands Member | Europe | ||
| Europe | |||
| Hrušky Formation | Europe | ||
| Gram Formation | Europe | ||
| Aquitaine Basin | Europe | ||
| Europe | |||
| Libano Sandstone | Europe | ||
| Blue Clay Formation | Europe | ||
| Globigerina Limestone[72] | Europe | ||
| Aalten Member | Europe | ||
| Breda Formation | Europe | ||
| Korytnica Clays | Europe | ||
| Leitha Limestone | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Filakovo Formation | Europe | ||
| Arjona Formation | Europe | ||
| Calcarenites of Sant Elm | Europe | ||
| Europe | |||
| Monterey Formation | North America | ||
| Puente Formation | North America | ||
| Purisima Formation | North America | ||
| San Mateo Formation | North America | ||
| Santa Margarita Formation | North America | ||
| Temblor Formation | North America | ||
| Topanga Formation | North America | ||
| Bone Valley Formation | North America | ||
| Calvert Formation | North America | ||
| Kirkwood Formation | North America | ||
| North America | |||
| Cojímar Formation | North America | ||
| Kendance Formation | North America | ||
| North America | |||
| Aymamón Limestone | North America | ||
| Almejas Formation | North America | ||
| Carrillo Puerto Formation | North America | ||
| Chagres Formation | North America | ||
| Chucunaque Formation | North America | ||
| Gatún Formation | North America | ||
| Paraná Formation | South America | ||
| Bahía Inglesa Formation | South America | ||
| Castilletes Formation | South America | ||
| Miramar Formation | South America | ||
| Pisco Formation | South America | ||
| Camacho Formation | South America | ||
| Cantaure Formation | South America | ||
| Caujarao Formation | South America | ||
| Socorro Formation | South America | ||
| Urumaco Formation | South America | ||
| Batesford Limestone | Oceania | ||
| Black Rock Sandstone | Oceania | ||
| Gippsland Limestone | Oceania | ||
| Mannum Formation | Oceania | ||
| Morgan Limestone | Oceania | ||
| Port Campbell Limestone | Oceania | ||
| Oceania | |||
| Oceania | |||
| Duho Formation[73] | Asia | ||
| Seogwipo Formation[74] | Asia |
Locations of megalodon fossil discoveries, yellow from the Pliocene and blue from the Miocene[26][71]
Prey relationships
Vertebra of a whale bitten in half by a megalodon with visible gashes from teeth.
Though sharks are generally opportunistic feeders, megalodon’s great size, high-speed swimming capability, and powerful jaws, coupled with an impressive feeding apparatus, made it an apex predator capable of consuming a broad spectrum of animals. Otodus megalodon was probably one of the most powerful predators to have existed.[15] A study focusing on calcium isotopes of extinct and extant elasmobranch sharks and rays revealed that megalodon fed at a higher trophic level than the contemporaneous great white shark («higher up» in the food chain.)[75]
Fossil evidence indicates that megalodon preyed upon many cetacean species, such as dolphins, small whales, cetotheres, squalodontids (shark toothed dolphins), sperm whales, bowhead whales, and rorquals.[51][76][77] In addition to this, they also targeted seals, sirenians, and sea turtles.[70] The shark was an opportunist and piscivorous, and it would have also gone after smaller fish and other sharks.[51] Many whale bones have been found with deep gashes most likely made by their teeth.[30]: 75 Various excavations have revealed megalodon teeth lying close to the chewed remains of whales,[30]: 75 [32] and sometimes in direct association with them.[78]
The feeding ecology of megalodon appears to have varied with age and between sites, like the modern great white shark. It is plausible that the adult megalodon population off the coast of Peru targeted primarily cetothere whales 2.5 to 7 meters (8.2 to 23 ft) in length and other prey smaller than itself, rather than large whales in the same size class as themselves.[76] Meanwhile, juveniles likely had a diet that consisted more of fish.[36][79]
Competition
Megalodon faced a highly competitive environment.[80] Its position at the top of the food chain[81] probably had a significant impact on the structuring of marine communities.[80][82] Fossil evidence indicates a correlation between megalodon and the emergence and diversification of cetaceans and other marine mammals.[30]: 78 [80] Juvenile megalodon preferred habitats where small cetaceans were abundant, and adult megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.[30]: 74–75
Megalodon were contemporaneous with whale-eating toothed whales (particularly macroraptorial sperm whales and squalodontidae), which were also probably among the era’s apex predators, and provided competition.[80] Some attained gigantic sizes, such as Livyatan, estimated between 13.5 to 17.5 meters (44 to 57 ft). Fossilized teeth of an undetermined species of such physeteroids from Lee Creek Mine, North Carolina, indicate it had a maximum body length of 8–10 m and a maximum lifespan of about 25 years. This is very different from similarly sized modern killer whales that live to 65 years, suggesting that unlike the latter, which are apex predators, these physeteroids were subject to predation from larger species such as megalodon or Livyatan.[83] By the Late Miocene, around 11 Mya, macroraptorials experienced a significant decline in abundance and diversity. Other species may have filled this niche in the Pliocene,[80][84] such as the fossil killer whale Orcinus citoniensis which may have been a pack predator and targeted prey larger than itself,[32][85][86][87] but this inference is disputed,[28] and it was probably a generalist predator rather than a marine mammal specialist.[88]
Megalodon may have subjected contemporaneous white sharks to competitive exclusion, as the fossil records indicate that other shark species avoided regions it inhabited by mainly keeping to the colder waters of the time.[89][30]: 77 In areas where their ranges seemed to have overlapped, such as in Pliocene Baja California, it is possible that megalodon and the great white shark occupied the area at different times of the year while following different migratory prey.[30]: 77 [90] Megalodon probably also had a tendency for cannibalism, much like contemporary sharks.[91]
Feeding strategies
Sharks often employ complex hunting strategies to engage large prey animals. Great white shark hunting strategies may be similar to how megalodon hunted its large prey.[92] Megalodon bite marks on whale fossils suggest that it employed different hunting strategies against large prey than the great white shark.[51]
One particular specimen–the remains of a 9-meter (30 ft) long undescribed Miocene baleen whale–provided the first opportunity to quantitatively analyze its attack behavior. Unlike great whites which target the underbelly of their prey, megalodon probably targeted the heart and lungs, with their thick teeth adapted for biting through tough bone, as indicated by bite marks inflicted to the rib cage and other tough bony areas on whale remains.[51] Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans, for example cetotheres, suggest that they were rammed with great force from below before being killed and eaten, based on compression fractures.[92]
There is also evidence that a possible separate hunting strategy existed for attacking raptorial sperm whales; a tooth belonging to an undetermined 4 m (13 ft) physeteroid closely resembling those of Acrophyseter discovered in the Nutrien Aurora Phosphate Mine in North Carolina suggests that a megalodon or O. chubutensis may have aimed for the head of the sperm whale in order to inflict a fatal bite, the resulting attack leaving distinctive bite marks on the tooth. While scavenging behavior cannot be ruled out as a possibility, the placement of the bite marks is more consistent with predatory attacks than feeding by scavenging, as the jaw is not a particularly nutritious area to for a shark feed or focus on. The fact that the bite marks were found on the tooth’s roots further suggest that the shark broke the whale’s jaw during the bite, suggesting the bite was extremely powerful. The fossil is also notable as it stands as the first known instance of an antagonistic interaction between a sperm whale and an otodontid shark recorded in the fossil record.[93]
During the Pliocene, larger cetaceans appeared.[94] Megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones and tail vertebrae of large whales from the Pliocene have been found with megalodon bite marks, which suggests that megalodon would immobilize a large whale before killing and feeding on it.[14][51]
Growth and reproduction
In 2010, Ehret estimated that megalodon had a fast growth rate nearly two times that of the extant great white shark. He also estimated that the slowing or cessation of somatic growth in megalodon occurred around 25 years of age, suggesting that this species had an extremely delayed sexual maturity.[95] In 2021, Shimada and colleagues calculated the growth rate of an approximately 9.2 m (30 ft) individual based on the Belgian vertebrate column specimen that presumably contains annual growth rings on three of its vertebrae. They estimated the individual died at 46 years of age, with a growth rate of 16 cm (6.3 in) per year, and a length of 2 m (6 ft 7 in) at birth. For a 15 m (49 ft) individual—which they considered to have been the maximum size attainable—this would equate to a lifespan of 88 to 100 years.[96] However, Cooper and his colleagues in 2022 estimated the length of this 46 year old individual at nearly 16 m (52 ft) based on the 3D reconstruction which resulted in the complete vertebral column to be 11.1 m (36 ft) long; the researchers claimed that this size estimate difference occurred due to the fact that Shimada and his colleagues extrapolated its size only based on the vertebral centra.[15]
Megalodon, like contemporaneous sharks, made use of nursery areas to birth their young in, specifically warm-water coastal environments with large amounts of food and protection from predators.[36] Nursery sites were identified in the Gatún Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands,[97] and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of megalodon also.[98] Infant megalodons were around 3.5 meters (11 ft) at their smallest,[40]: 61 and the pups were vulnerable to predation by other shark species, such as the great hammerhead shark (Sphyrna mokarran) and the snaggletooth shark (Hemipristis serra).[36] Their dietary preferences display an ontogenetic shift:[40]: 65 Young megalodon commonly preyed on fish,[36] sea turtles,[70] dugongs,[31]: 129 and small cetaceans; mature megalodon moved to off-shore areas and consumed large cetaceans.[30]: 74–75
An exceptional case in the fossil record suggests that juvenile megalodon may have occasionally attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-to-7-meter (13 to 23 ft) long Pliocene shark were found on a rib from an ancestral blue or humpback whale that showed evidence of subsequent healing, which is suspected to have been inflicted by a juvenile megalodon.[99][100]
Extinction
Climate change
The Earth experienced a number of changes during the time period megalodon existed which affected marine life. A cooling trend starting in the Oligocene 35 Mya ultimately led to glaciation at the poles. Geological events changed currents and precipitation; among these were the closure of the Central American Seaway and changes in the Tethys Ocean, contributing to the cooling of the oceans. The stalling of the Gulf Stream prevented nutrient-rich water from reaching major marine ecosystems, which may have negatively affected its food sources. The largest fluctuation of sea levels in the Cenozoic era occurred in the Plio-Pleistocene, between around 5 million to 12 thousand years ago, due to the expansion of glaciers at the poles, which negatively impacted coastal environments, and may have contributed to its extinction along with those of several other marine megafaunal species.[101] These oceanographic changes, in particular the sea level drops, may have restricted many of the suitable shallow warm-water nursery sites for megalodon, hindering reproduction.[102] Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.[103][36]
As its range did not apparently extend into colder waters, megalodon may not have been able to retain a significant amount of metabolic heat, so its range was restricted to shrinking warmer waters.[102][77][104] Fossil evidence confirms the absence of megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[30]: 77 However, an analysis of the distribution of megalodon over time suggests that temperature change did not play a direct role in its extinction. Its distribution during the Miocene and Pliocene did not correlate with warming and cooling trends; while abundance and distribution declined during the Pliocene, megalodon did show a capacity to inhabit colder latitudes. It was found in locations with a mean temperature ranging from 12 to 27 °C (54 to 81 °F), with a total range of 1 to 33 °C (34 to 91 °F), indicating that the global extent of suitable habitat should not have been greatly affected by the temperature changes that occurred.[26] This is consistent with evidence that it was a mesotherm.[49]
Changing ecosystem
Marine mammals attained their greatest diversity during the Miocene,[30]: 71 such as with baleen whales with over 20 recognized Miocene genera in comparison to only six extant genera.[106] Such diversity presented an ideal setting to support a super-predator such as megalodon.[30]: 75 By the end of the Miocene, many species of mysticetes had gone extinct;[80] surviving species may have been faster swimmers and thus more elusive prey.[31]: 46 Furthermore, after the closure of the Central American Seaway, tropical whales decreased in diversity and abundance.[104] The extinction of megalodon correlates with the decline of many small mysticete lineages, and it is possible that it was quite dependent on them as a food source.[76] Additionally, a marine megafauna extinction during the Pliocene was discovered to have eliminated 36% of all large marine species including 55% of marine mammals, 35% of seabirds, 9% of sharks, and 43% of sea turtles. The extinction was selective for endotherms and mesotherms relative to poikilotherms, implying causation by a decreased food supply[101] and thus consistent with megalodon being mesothermic.[49] Megalodon may have been too large to sustain itself on the declining marine food resources.[102] The cooling of the oceans during the Pliocene might have restricted the access of megalodon to the polar regions, depriving it of the large whales which had migrated there.[104]
Competition from large odontocetes, such as macropredatory sperm whales which appeared in the Miocene, and a member of genus Orcinus (i.e., Orcinus citoniensis) in the Pliocene,[80][84] is assumed to have contributed to the decline and extinction of megalodon.[26][31]: 46–47 [102][107] But this assumption is disputed:[28] The Orcininae emerged in Mid-Pliocene with O. citoniensis reported from the
Pliocene of Italy,[84][108] and similar forms reported from the Pliocene of England and South Africa,[84] indicating the capacity of these dolphins to cope with increasingly prevalent cold water temperatures in high latitudes.[84] These dolphins were assumed to have been macrophagous in some studies,[26] but on closer inspection, these dolphins are not found to be macrophagous and fed on small fishes instead.[108] On the other hand, gigantic macropredatory sperm whales such as Livyatan-like forms are last reported from Australia and South Africa circa 5 million years ago.[109][110][111] Others, such as Hoplocetus and Scaldicetus also occupied a niche similar to that of modern killer whales but the last of these forms disappeared during the Pliocene.[112][108] Members of genus Orcinus became large and macrophagous in the Pleistocene.[108]
Paleontologist Robert Boessenecker and his colleagues rechecked the fossil record of megalodon for carbon dating errors and concluded that it disappeared circa 3.5 million years ago.[28] Boessenecker and his colleagues further suggest that megalodon suffered range fragmentation due to climatic shifts,[28] and competition with white sharks might have contributed to its decline and extinction.[28] Competition with white sharks is assumed to be a factor in other studies as well,[113][26][107] but this hypothesis warrants further testing.[114] Multiple compounding environmental and ecological factors including climate change and thermal limitations, collapse of prey populations and resource competition with white sharks are believed to have contributed to decline and extinction of megalodon for now.[107]
The extinction of megalodon set the stage for further changes in marine communities. The average body size of baleen whales increased significantly after its disappearance, although possibly due to other, climate-related, causes.[115] Conversely the increase in baleen whale size may have contributed to the extinction of megalodon, as they may have preferred to go after smaller whales; bite marks on large whale species may have come from scavenging sharks. Megalodon may have simply become coextinct with smaller whale species, such as Piscobalaena nana.[105] The extinction of megalodon had a positive impact on other apex predators of the time, such as the great white shark, in some cases spreading to regions where megalodon became absent.[26][113][116]
In popular culture
HMS Challenger discovered megalodon teeth which were erroneously dated to be around 11,000 to 24,000 years old.
Megalodon has been portrayed in many works of fiction, including films and novels, and continues to be a popular subject for fiction involving sea monsters.[117] Reports of supposedly fresh megalodon teeth, such as those found by HMS Challenger in 1873 which were dated in 1959 by the zoologist Wladimir Tschernezky to be around 11,000 to 24,000 years old, helped popularise claims of recent megalodon survival amongst cryptozoologists.[118] These claims are now discredited, and are probably teeth that were well-preserved by a thick mineral-crust precipitate of manganese dioxide, and so had a lower decomposition rate and retained a white color during fossilization. Fossil megalodon teeth can vary in color from off-white to dark browns and greys, and some fossil teeth may have been redeposited into a younger stratum. The claims that megalodon could remain elusive in the depths, similar to the megamouth shark which was discovered in 1976, are unlikely as the shark lived in warm coastal waters and probably could not survive in the cold and nutrient-poor deep sea environment.[119][120]
Contemporary fiction about megalodon surviving into modern times was pioneered by the 1997 novel Meg: A Novel of Deep Terror by Steve Alten and its subsequent sequels. Megalodon subsequently began to feature in films, such as the 2003 direct to video Shark Attack 3: Megalodon, and later The Meg, a 2018 film based on the 1997 book which grossed over $500 million at the box office.[118][121]
Animal Planet’s pseudo-documentary Mermaids: The Body Found included an encounter 1.6 mya between a pod of mermaids and a megalodon.[122] Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives,[123] a controversial docufiction about the creature that presented alleged evidence in order to suggest that megalodons still lived. This program received criticism for being completely fictional and for inadequately disclosing its fictional nature; for example, all of the supposed scientists depicted were paid actors, and there was no disclosure in the documentary itself that it was fictional. In a poll by Discovery, 73% of the viewers of the documentary thought that megalodon was not extinct. In 2014, Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.[51][124][125][126] Despite the criticism from scientists, Megalodon: The Monster Shark Lives was a huge ratings success, gaining 4.8 million viewers, the most for any Shark Week episode up to that point.[127]
Megalodon teeth are the state fossil of North Carolina.[128]
See also
- List of prehistoric cartilaginous fish
- Prehistoric fish
- Largest prehistoric organisms
Notes
- ^ Carbonated bioapatite from a megalodon tooth (of unknown source location) dated to 5.75 ± 0.9 Ma in age has been analyzed for isotope ratios of oxygen (18O/16O) and carbon (13C/12C), using a carbonate clumped-isotope thermometer methodology to yield an estimate of the ambient temperature in that individual’s environment of 19 ± 4 °C.[69]
References
- ^ a b Agassiz, Louis (1843). Recherches sur les poissons fossiles [Research on the fossil fishes] (in French). Neuchatel: Petitpierre. p. 41.
- ^ «Otodus (Megaselachus) megalodon (Agassiz, 1837)». SharkReferences.com. Retrieved 24 October 2017.
- ^ Eastman, C. R. (1904). Maryland Geological Survey. Vol. 2. Baltimore, Maryland: Johns Hopkins University. p. 82.
- ^ a b c Cappetta, H. (1987). «Mesozoic and Cenozoic Elasmobranchii». Handbook of Paleoichthyology. Vol. 3B. München, Germany: Friedrich Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- ^ Hay, O. P. (1901). «Bibliography and Catalogue of the Fossil Vertebrata of North America». Bulletin of the United States Geological Society (179): 308.
- ^ a b c d e f Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (2016). «A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade». Historical Biology. 29 (5): 1–11. doi:10.1080/08912963.2016.1236795. S2CID 89080495.
- ^ a b c d e f Shimada, Kenshu (2019). «The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited». Historical Biology. 33 (7): 1–8. doi:10.1080/08912963.2019.1666840. ISSN 0891-2963. S2CID 208570844.
- ^ a b Cooper, J. A.; Pimiento, C.; Ferrón, H. G.; Benton, M. J. (2020). «Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction». Scientific Reports. 10 (14596): 14596. Bibcode:2020NatSR..1014596C. doi:10.1038/s41598-020-71387-y. PMC 7471939. PMID 32883981.
- ^ «Giant ‘megalodon’ shark extinct earlier than previously thought». Science Daily. 13 February 2019.
- ^ a b c d e Perez, Victor; Leder, Ronny; Badaut, Teddy (2021). «Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions». Palaeontologia Electronica. 24 (1): 1–28. doi:10.26879/1140.
- ^ a b Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c d e Pimiento, C.; Balk, M. A. (2015). «Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators». Paleobiology. 41 (3): 479–490. doi:10.1017/pab.2015.16. PMC 4541548. PMID 26321775.
- ^ a b c Prothero, Donald R. (25 August 2015). «09. Mega-Jaws: The Largest Fish. Carcharocles». The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution. New York Chichester, West Sussex: Columbia University Press. pp. 96–110. doi:10.7312/prot17190-010. ISBN 978-0-231-53942-5.
- ^ a b c d e Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?» (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ^ a b c d e f Cooper, J.A.; Hutchinson, J.R.; Bernvi, D.C.; Cliff, G.; Wilson, R.P.; Dicken, M.L.; Menzel, J.; Wroe, S.; Pirlo, J.; Pimiento, C. (2022). «The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling». Science Advances. 8 (33): eabm9424. Bibcode:2022SciA….8M9424C. doi:10.1126/sciadv.abm9424. ISSN 2375-2548. PMC 9385135. PMID 35977007.
- ^ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Westport, Connecticut: Libraries Unlimited. pp. 25–26. ISBN 978-1-59158-265-6. OCLC 230807846.
- ^ Hsu, Kuang-Tai (2009). «The Path to Steno’s Synthesis on the Animal Origin of Glossopetrae». In Rosenburg, G. D. (ed.). The Revolution in Geology from the Renaissance to the Enlightenment. Vol. 203. Boulder, Colorado: Geological Society of America. ISBN 978-0-8137-1203-1. OCLC 608657795.
- ^ Eilperin, J. (2012). Demon Fish. Pantheon Books. p. 43. ISBN 978-0-7156-4352-5.
- ^ a b c d e f Nyberg, K. G.; Ciampaglio C. N.; Wray G. A. (2006). «Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth». Journal of Vertebrate Paleontology. 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. S2CID 53640614.
- ^ Keyes, I. W. (2012). «New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record». New Zealand Journal of Geology and Geophysics. 15 (2): 229. doi:10.1080/00288306.1972.10421956.
- ^ μέγας. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ ὀδούς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ Lawley, R. (1881). «Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)» [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (in Italian). 5: 167–172.
- ^ Yabe, H.; Goto, M.; Kaneko, N. (2004). «Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records». The Palaeontological Society of Japan. 75: 7–15.
- ^ a b c Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ a b c d e f g h i j k l Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c Pimiento, C.; Clements, C. F. (2014). «When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record». PLOS ONE. 9 (10): e111086. Bibcode:2014PLoSO…9k1086P. doi:10.1371/journal.pone.0111086. PMC 4206505. PMID 25338197.
- ^ a b c d e f Boessenecker, R. W.; Ehret, D. J.; Long, D. J.; Churchill, M.; Martin, E.; Boessenecker, S. J. (2019). «The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific». PeerJ. 7: e6088. doi:10.7717/peerj.6088. PMC 6377595. PMID 30783558.
- ^ a b c d Ehret D. J.; Hubbell G.; Macfadden B. J. (2009). «Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru». Journal of Vertebrate Paleontology. 29 (1): 1–13. doi:10.1671/039.029.0113. JSTOR 20491064. S2CID 129585445.
- ^ a b c d e f g h i j k l m n o p q r Klimley, Peter; Ainley, David (1996). «Evolution». Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. ISBN 978-0-12-415031-7. OCLC 212425118.
- ^ a b c d e f g h i j k l m n o p Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ a b c d e Andres, Lutz (2002). «C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles«. fossilguy.com. Retrieved 16 January 2008.
- ^ a b Perez, V. J.; Godfrey, S. J.; Kent, B. W.; Weems, R. E.; Nance, J. R. (2019). «The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time». Journal of Vertebrate Paleontology. 38 (6): e1546732. doi:10.1080/02724634.2018.1546732.
- ^ Siverson, M.; Lindgren, J.; Newbrey, M.G.; Cederström, P.; Cook, T.D. (2013). «Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type» (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. S2CID 58906204. Archived from the original (PDF) on 19 October 2013.
- ^ Benton, M. J.; Pearson, P. N. (2001). «Speciation in the fossil record». Trends in Ecology and Evolution. 16 (7): 405–411. doi:10.1016/s0169-5347(01)02149-8. PMID 11403874.
- ^ a b c d e f g h Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Vivian, G. (2013). «Research debunks Great White lineage».
- ^ «Palaeontological detective work unravels evolution of megatooth sharks». 2013.
- ^ a b c d e f g h i j k l m n o p q r Gottfried, MD; Compagno, LJV; Bowman, SC (1996). «Size and skeletal anatomy of the giant megatooth shark Carcharodon megalodon«. In Klimley; Ainley (eds.). Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. pp. 55–89. ISBN 978-0124150317.
- ^ «Could Megalodon Have Looked Like a BIG Sandtiger Shark?». Biology of Sharks and Rays. Retrieved 2 September 2017.
- ^ Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). «Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles» (PDF). Caribbean Journal of Science. 44 (3): 279–286. doi:10.18475/cjos.v44i3.a2. S2CID 87154947. Archived from the original (PDF) on 20 July 2011.
- ^ Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ a b McClain, Craig R.; Balk, Meghan A.; Benfield, Mark C.; Branch, Trevor A.; Chen, Catherine; Cosgrove, James; Dove, Alistair D.M.; Gaskins, Lindsay C.; Helm, Rebecca R.; Hochberg, Frederick G.; Lee, Frank B. (13 January 2015). «Sizing ocean giants: patterns of intraspecific size variation in marine megafauna». PeerJ. 3: e715. doi:10.7717/peerj.715. ISSN 2167-8359. PMC 4304853. PMID 25649000.
- ^ I., Castro, Jose (2011). Sharks of North America. Oxford University Press, USA. ISBN 978-0-19-978097-6. OCLC 958576172.
- ^ Borrell, Asunción; Aguilar, Alex; Gazo, Manel; Kumarran, R. P.; Cardona, Luis (1 December 2011). «Stable isotope profiles in whale shark (Rhincodon typus) suggest segregation and dissimilarities in the diet depending on sex and size». Environmental Biology of Fishes. 92 (4): 559–567. doi:10.1007/s10641-011-9879-y. ISSN 1573-5133. S2CID 37683420.
- ^ Jacoby, D. M. P.; Siriwat, P.; Freeman, R.; Carbone, C. (2015). «Is the scaling of swim speed in sharks driven by metabolism?». Biology Letters. 12 (10): 20150781. doi:10.1098/rsbl.2015.0781. PMC 4707698. PMID 26631246.
- ^ a b c Ferrón, H. G. (2017). «Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks». PLOS ONE. 12 (9): e0185185. Bibcode:2017PLoSO..1285185F. doi:10.1371/journal.pone.0185185. PMC 5609766. PMID 28938002.
- ^ Shimada, K.; Becker, M. A.; Griffiths, M. L. (2020). «Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to ‘off-the-scale’ gigantism of the megatooth shark, Otodus megalodon«. Historical Biology. 33 (11): 1–17. doi:10.1080/08912963.2020.1812598.
- ^ a b c d e f g Prothero, D. R. (2015). «Mega-Jaws». The Story of Life in 25 Fossils. New York, New York: Columbia University Press. pp. 96–110. ISBN 978-0-231-17190-8. OCLC 897505111.
- ^ Helfman, G.; Burgess, G. H. (2014). Sharks: The Animal Answer Guide. Baltimore, Maryland: Johns Hopkins University Press. p. 19. ISBN 978-1-4214-1310-5. OCLC 903293986.
- ^ Randall, John E. (1973). «Size of the Great White Shark (Carcharodon)». Science Magazine. 181 (4095): 169–170. Bibcode:1973Sci…181..169R. doi:10.1126/science.181.4095.169. PMID 17746627. S2CID 36607712.
- ^ Schembri, Patrick (1994). «Malta’s Natural Heritage». Natural Heritage. In: 105–124.
- ^ Papson, Stephen (1992). «Copyright: Cross the Fin Line of Terror». Journal of American Culture. 15 (4): 67–81. doi:10.1111/j.1542-734X.1992.1504_67.x.
- ^ Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ Shimada, Kenshu (2002). «The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)». Journal of Fossil Research. 35 (2): 28–33.
- ^ a b Pimiento, Catalina; Gerardo González-Barba; Dana J. Ehret; Austin J. W. Hendy; Bruce J. MacFadden; Carlos Jaramillo (2013). «Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama» (PDF). Journal of Paleontology. 87 (5): 755–774. doi:10.1666/12-117. S2CID 45662900. Archived from the original (PDF) on 29 October 2013.
- ^ Crane, B. (2017). «A Prehistoric Killer, Buried in Muck». The New Yorker. Retrieved 10 December 2017.
- ^ Mustain, A. (2011). «For Sale: World’s Largest Shark Jaws». LiveScience. Retrieved 31 August 2017.
- ^ a b Almgreen, S. E. Bendix (15 November 1983). «Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn». Bulletin of the Geological Society of Denmark. 32: 1–32. CiteSeerX 10.1.1.514.1782. doi:10.37570/bgsd-1983-32-01. S2CID 53311833. NAID 10012345550.
- ^ Reolid, M.; Molina, J. M. (2015). «Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain)». Estudios Geológicos. 71 (2): e032. doi:10.3989/egeol.41828.342.
- ^ Uyeno, T.; Sakamoto, O.; Sekine, H. (1989). «The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan». Saitama Museum of Natural History Bulletin. 7: 73–85.
- ^ Anderson, P.S.L.; Westneat, M. (2009). «A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)». Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1. S2CID 86203770.
- ^ Ballell, A.; Ferrón, H.G. (2021). «Biomechanical insights into the dentition of megatooth sharks (Lamniformes: Otodontidae)». Scientific Reports. 11 (1232): 1232. doi:10.1038/s41598-020-80323-z. PMC 7806677. PMID 33441828.
- ^ a b Stringer, G. L.; King, L. (2012). «Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana». New Mexico Museum of Natural History and Science, Bulletin. Vertebrate Corpolites. 57: 301.
- ^ «Megalodon Shark Facts and Information: The Details». fossilguy.com. Retrieved 18 September 2017.
- ^ Fitzgerald, Erich (2004). «A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia». Memoirs of Museum Victoria. 61 (2): 183–208. doi:10.24199/j.mmv.2004.61.12.
- ^ Löffler, N.; Fiebig, J.; Mulch, A.; Tütken, T.; Schmidt, B.C.; Bajnai, D.; Conrad, A.C.; Wacker, U.; Böttcher, M.E. (2019). «Refining the temperature dependence of the oxygen and clumped isotopic compositions of structurally bound carbonate in apatite». Geochimica et Cosmochimica Acta. 253: 19–38. Bibcode:2019GeCoA.253…19L. doi:10.1016/j.gca.2019.03.002. S2CID 107992832.
- ^ a b c Aguilera O.; Augilera E. R. D. (2004). «Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage». Caribbean Journal of Science. 40 (3): 362–368.
- ^ a b «Carcharocles megalodon«. Fossilworks. Retrieved 28 August 2017 from the Paleobiology Database.
{{cite web}}: CS1 maint: postscript (link) - ^ «Young Prince George gifted 23m-year-old tooth from extinct shark found in Malta». Times of Malta. 27 September 2020. Retrieved 28 September 2020.
- ^ Yun, C. (2020). «New example of Cosmopolitodus hastalis (Lamniformes, Lamnidae) from the Miocene South Korea». Zoodiversity. 54 (5): 433–438. doi:10.15407/zoo2020.05.433. S2CID 229274996.
- ^ Choi, Seung; Lee, Yuong-Nam (2017). «A review of vertebrate body fossils from the Korean Peninsula and perspectives». Geosciences Journal. 21 (6): 867–889. Bibcode:2017GescJ..21..867C. doi:10.1007/s12303-017-0040-6. ISSN 1226-4806. S2CID 133835817.
- ^ Martin, J. E.; Tacail, T.; Sylvain, A.; Catherine, G.; Vincent, B. (2015). «Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs». Chemical Geology. 415: 118–125. Bibcode:2015ChGeo.415..118M. doi:10.1016/j.chemgeo.2015.09.011.
- ^ a b c Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ a b Morgan, Gary S. (1994). «Whither the giant white shark?» (PDF). Paleontology Topics. 2 (3): 1–2. Archived from the original (PDF) on 22 July 2016.
- ^ Augilera, Orangel A.; García, Luis; Cozzuol, Mario A. (2008). «Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation». Paläontologische Zeitschrift. 82 (2): 204–208. doi:10.1007/BF02988410. ISSN 0038-2353. S2CID 84251638.
- ^ Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). «The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)». Journal of South American Earth Sciences. 73: 168–190. Bibcode:2017JSAES..73..168L. doi:10.1016/j.jsames.2016.12.010.
- ^ a b c d e f g h Lambert, O.; Bianucci, G.; Post, P.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. (2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020. S2CID 4369352.
- ^ Compagno, Leonard J. V. (1989). «Alternative life-history styles of cartilaginous fishes in time and space». Environmental Biology of Fishes. 28 (1–4): 33–75. doi:10.1007/BF00751027. S2CID 22527888.
- ^ Ferretti, Francesco; Boris Worm; Gregory L. Britten; Michael R. Heithaus; Heike K. Lotze1 (2010). «Patterns and ecosystem consequences of shark declines in the ocean» (PDF). Ecology Letters. 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. Archived from the original (PDF) on 6 July 2011. Retrieved 19 February 2011.
- ^ Gilbert, K.N.; Ivany, L.C.; Uhen, M.D. (2018). «Living fast and dying young: life history and ecology of a Neogene sperm whale». Journal of Vertebrate Paleontology. 38 (2): e1439038. doi:10.1080/02724634.2018.1439038. S2CID 89750852.
- ^ a b c d e Heyning, John; Dahlheim, Marilyn (1988). «Orcinus orca» (PDF). Mammalian Species (304): 1–9. doi:10.2307/3504225. JSTOR 3504225. S2CID 253914153. Archived from the original (PDF) on 5 December 2010.
- ^ Bianucci, Giovanni; Walter, Landini (2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society. 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x.
- ^ Lindberg, D. R.; Pyenson, N. D. (2006). «Evolutionary Patterns in Cetacea: Fishing Up Prey Size through Deep Time». Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 77. ISBN 978-0-520-24884-7.
- ^ Boessenecker, R. W. (2013). «A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans». Geodiversitas. 35 (4): 815–940. doi:10.5252/g2013n4a5. S2CID 85940452.
- ^ Bianucci, G. (1997). «Hemisyntrachelus cortesii (Cetacea, Delphinidae) from the Pliocene Sediments of Campore Quarry (Salsomaggiori Terme, Italy». Bollettino della Societa Paleontologica Italiana. 36 (1): 75–83).
- ^ Antunes, M.T.; Legoinha, P.; Balbing, A. (2015). «Megalodon, mako shark and planktonic foraminifera from the continental shelf off Portugal and their age». Geologica Acta. 13: 181–190.
- ^ «Paleoecology of Megalodon and the White Shark». Biology of Sharks and Rays. Retrieved 1 October 2017.
- ^ Tanke, Darren; Currie, Philip (1998). «Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence» (PDF). Gaia (15): 167–184.
- ^ a b Godfrey, S. J.; Altman, J. (2005). «A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon» (PDF). Jeffersoniana (16): 1–12.
- ^ STEPHEN J. GODFREY; JOHN R. NANCE; NORMAN L. RIKER (2021). «Otodus-bitten sperm whale tooth from the Neogene of the Coastal Eastern United States» (PDF). Acta Palaeontologica Polonica. 66 (3): 599–603.
- ^ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). «The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes». Journal of Mammalian Evolution. 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3. S2CID 90231.
- ^ Ehret D.J. (2010). «CHAPTER 5 – MACROEVOLUTION, AGE, AND GROWTH DETERMINATION OF THE MEGATOOTHED SHARKS (LAMNIFORMES: OTODONTIDAE)». Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes) (PDF). pp. 100–136.
- ^ S., Kenshu; Bonnan, M. F.; Becker, M. A.; Griffiths, M. L. (2021). «Ontogenetic growth pattern of the extinct megatooth shark Otodus megalodon—implications for its reproductive biology, development, and life expectancy». Historical Biology. 33 (12): 3254–3259. doi:10.1080/08912963.2020.1861608.
- ^ «Identifican en Canarias fósiles de ‘megalodón’, el tiburón más grande que ha existido» [Identifying Canary fossils of ‘megalodon’, the largest shark that ever lived] (in Spanish). Europa Press Noticias SA. 2013. Retrieved 29 August 2017.
- ^ Dulvy, N. K.; Reynolds, J. D. (1997). «Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays». Proceedings of the Royal Society B: Biological Sciences. 264 (1386): 1309–1315. Bibcode:1997RSPSB.264.1309D. doi:10.1098/rspb.1997.0181. PMC 1688595.
- ^ Godfrey, Stephen (2004). «The Ecphora» (PDF). The Newsletter of Calvert Marine Museum Fossil Club. 19 (1): 1–13. Archived from the original (PDF) on 10 December 2010.
- ^ Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (27 August 2010). «Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event». International Journal of Osteoarchaeology. 22 (3): 253–260. doi:10.1002/oa.1199.
- ^ a b Pimiento, C.; Griffin, J. N.; Clements, C. F.; Silvestro, D.; Varela, S.; Uhen, M. D.; Jaramillo, C. (2017). «The Pleistocene Marine Megafauna Extinction and its Impact on Functional Diversity». Nature Ecology and Evolution. 1 (8): 1100–1106. doi:10.1038/s41559-017-0223-6. PMID 29046566. S2CID 3639394.
- ^ a b c d «The Extinction of Megalodon». Biology of Sharks and Rays. Retrieved 31 August 2017.
- ^ Reilly, Michael (29 September 2009). «Prehistoric Shark Nursery Spawned Giants». Discovery News. Archived from the original on 10 March 2012. Retrieved 23 November 2013.
- ^ a b c Allmon, Warren D.; Steven D. Emslie; Douglas S. Jones; Gary S. Morgan (2006). «Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms». The Journal of Geology. 104 (2): 143–162. Bibcode:1996JG….104..143A. doi:10.1086/629811. S2CID 128418299.
- ^ a b Collareta, A.; Lambert, O.; Landini, W.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ Dooly A.C.; Nicholas C. F.; Luo Z. X. (2006). «The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)». Journal of Vertebrate Paleontology. 24 (2): 453–463. doi:10.1671/2401. JSTOR 4524731. S2CID 84970052.
- ^ a b c McCormack, Jeremy; Griffiths, Michael L.; Kim, Sora L.; Shimada, Kenshu; Karnes, Molly; Maisch, Harry; Pederzani, Sarah; Bourgon, Nicolas; Jaouen, Klervia; Becker, Martin A.; Jöns, Niels (31 May 2022). «Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes». Nature Communications. 13 (1): 2980. Bibcode:2022NatCo..13.2980M. doi:10.1038/s41467-022-30528-9. ISSN 2041-1723. PMC 9156768. PMID 35641494. S2CID 249235478.
- ^ a b c d Citron, Sara; Geisler, Jonathan H.; Alberto, Collareta; Giovanni, Bianucci (2022). «Systematics, phylogeny and feeding behavior of the oldest killer whale: a reappraisal of Orcinus citoniensis (Capellini, 1883) from the Pliocene of Tuscany (Italy)». Bollettino della Società Paleontologica Italiana. 61 (2): 167–186. doi:10.4435/BSPI.2022.13.
- ^ «Huge Tooth Reveals Prehistoric Moby Dick in Melbourne». Australasian Science Magazine. Retrieved 24 April 2016.
- ^ «Move over Moby Dick: Meet Melbourne’s own mega whale». The Sydney Morning Herald. 21 April 2016.
- ^ Govender, R (2021). «Early Pliocene fossil cetaceans from Hondeklip Bay, Namaqualand, South Africa». Historical Biology. 33 (4): 574–593. doi:10.1080/08912963.2019.1650273. S2CID 202019648.
- ^ Hampe, O. (2006). «Middle/late Miocene hoplocetine sperm whale remains (Odontoceti: Physeteridae) of North Germany with an emended classification of the Hoplocetinae». Fossil Record. 9 (1): 61–86. doi:10.1002/mmng.200600002.
- ^ a b Antunes, Miguel Telles; Balbino, Ausenda Cáceres (2010). «The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic». Revista Española de Paleontología. 25 (1): 1–6.
- ^ Kast, Emma R.; Griffiths, Michael L.; Kim, Sora. L.; Rao, Zixuan C.; Shimada, Kensu; Becker, Martin A.; Maisch, Harry M.; Eagle, Robert A.; Clarke, Chelesia A.; Neumann, Allison N.; Karnes, Molly E.; Lüdecke, Tina; Leichliter, Jennifer N.; Martínez-García, Alfredo; Akhtar, Alliya A.; Wang, Xingchen T.; Haug, Gerald H.; Sigman, Daniel M. (22 June 2022). «Cenozoic megatooth sharks occupied extremely high trophic positions». Science Advances. 8 (25): eabl6529. Bibcode:2022SciA….8L6529K. doi:10.1126/sciadv.abl6529. PMC 9217088. PMID 35731884.
- ^ Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (2017). «Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics». Proceedings of the Royal Society B: Biological Sciences. 284 (1855): 20170546. doi:10.1098/rspb.2017.0546. PMC 5454272. PMID 28539520.
- ^ Sylvain, Adnet; A. C. Balbino; M. T. Antunes; J. M. Marín-Ferrer (2010). «New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean». Neues Jahrbuch für Geologie und Paläontologie. 256 (1): 7–16. doi:10.1127/0077-7749/2009/0029.
- ^ Weinstock, J. A. (2014). The Ashgate Encyclopedia of Literary and Cinematic Monsters. Farnham, United Kingdom: Routledge. pp. 107–108. ISBN 978-1-4094-2562-5. OCLC 874390267.
- ^ a b Guimont, Edward (5 October 2021). «The Megalodon: A Monster of the New Mythology». M/C Journal. 24 (5). doi:10.5204/mcj.2793. ISSN 1441-2616. S2CID 241813307.
- ^ Roesch, B. S. (1998). «A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles megalodon«. The Cryptozoology Review. 3 (2): 14–24.
- ^ «Does Megalodon Still Live?». Biology of Sharks and Rays. Retrieved 2 October 2017.
- ^ «The Meg (2018)». Box Office Mojo. Retrieved 22 December 2017.
- ^ Sid Bennett (director) (27 May 2012). Mermaids: The Body Found (Motion picture). Animal Planet.
- ^ «Shark Week ‘Megalodon: The Monster Shark Lives’ Tries To Prove Existence Of Prehistoric Shark (VIDEO)». Huff Post Green. 5 August 2013. Retrieved 11 August 2013.
- ^ Winston, B.; Vanstone, G.; Chi, W. (2017). «A Walk in the Woods». The Act of Documenting: Documentary Film in the 21st Century. New York, New York: Bloomsbury Publishing. ISBN 978-1-5013-0918-2. OCLC 961183719.
- ^ Flanagin, J. (2014). «Sorry, Fans. Discovery Has Jumped the Shark Week». New York Times. Retrieved 16 August 2014.
- ^ Shiffman, David (15 August 2014). «Shark Week Is Lying Again About Monster Megalodon Sharks». Slate Magazine. Retrieved 31 July 2022.
- ^ O’Connell, Mikey (5 August 2013). «TV Ratings: Shark Week Hits Record Highs With Fake ‘Megalodon’ Doc». The Hollywood Reporter. Retrieved 31 July 2022.
- ^ «Fossil, Fossilized Teeth of the Megalodon Shark | NCpedia». ncpedia.org. Retrieved 17 October 2019.
Further reading
- Dickson, K. A.; Graham, J. B. (November–December 2004). «Evolution and consequences of endothermy in fishes». Physiological and Biochemical Zoology. 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772. S2CID 40104003.
- Kent, Bretton W. (1994). Fossil Sharks of the Chesapeake Bay Region. Columbia, Md.: Egan Rees & Boyer. ISBN 978-1-881620-01-3. OCLC 918266672.
External links
- The rise of super predatory sharks
- Carcharocles: Extinct Megatoothed shark
- Paleontological videos
- Paleontologist Mark Renz shows one of the largest megalodon teeth discovered on YouTube
- Shark Week Special on megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about megalodon on YouTube (featuring expert Dana Ehret)
- Lamniform sharks: 110 million years of ocean supremacy on YouTube (featuring expert Mikael Siverson)
- The Rise and Fall of the Neogene Giant Sharks on YouTube (featuring expert Bretton Kent)
- Moore, Kallie (19 December 2018). «Why Megalodon (Definitely) Went Extinct». PBS Eons. Archived from the original on 14 November 2021 – via YouTube.
| Megalodon
Temporal range: Early Miocene–Early Pliocene, c. 23–3.6 Ma PreꞒ Ꞓ O S D C P T J K Pg N |
|
|---|---|
|
|
|
| Model of megalodon jaws at the American Museum of Natural History | |
| Scientific classification |
|
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Otodontidae |
| Genus: | †Otodus |
| Species: |
†O. megalodon |
| Binomial name | |
| †Otodus megalodon
(Agassiz, 1843)[1] |
|
| Synonyms[2][3][4][5][6] | |
|
List of synonyms
|
Megalodon (Otodus megalodon),[6][7][8] meaning «big tooth», is an extinct species of mackerel shark that lived approximately 23 to 3.6 million years ago (Mya), from the Early Miocene to the Pliocene epochs.[9] It was formerly thought to be a member of the family Lamnidae and a close relative of the great white shark (Carcharodon carcharias). However, it is now classified into the extinct family Otodontidae, which diverged from the great white shark during the Early Cretaceous.
While regarded as one of the largest and most powerful predators to have ever lived, the megalodon is only known from fragmentary remains, and its appearance and maximum size are uncertain. Scientists differ on whether it would have more closely resembled a stockier version of the great white shark, the whale shark (Rhincodon typus), the basking shark (Cetorhinus maximus) or the sand tiger shark (Carcharias taurus). The most recent estimate with the least error range suggests a maximum length estimate up to 20.3 meters (67 ft),[10] although the modal lengths are estimated at 10.5 meters (34 ft).[11][12] Extrapolation from a vertebral centra with dimensions based on the great white shark suggests that a megalodon about 16 meters (52 ft) long weighs up to 48 metric tons (53 short tons), 17 meters (56 ft) long weighs up to 59 metric tons (65 short tons), and 20.3 meters (67 ft) long (the maximum length) weighs up to 103 metric tons (114 short tons).[13][14] Extrapolating from a vertebral column and reconstructing a 3D model with dimensions based on all extant lamnid sharks suggests that a 16-meter-long (52 ft) individual may have been much larger than previous estimates, reaching an excess of 61.5 metric tons (67.8 short tons) in body mass; an individual of this size would have needed to consume 98,175 kcal per day.[15] Their teeth were thick and robust, built for grabbing prey and breaking bone, and their large jaws could exert a bite force of up to 108,500 to 182,200 newtons (24,400 to 41,000 lbf).[14]
Megalodon probably had a major impact on the structure of marine communities. The fossil record indicates that it had a cosmopolitan distribution. It probably targeted large prey, such as whales, seals and sea turtles. Juveniles inhabited warm coastal waters and fed on fish and small whales. Unlike the great white, which attacks prey from the soft underside, megalodon probably used its strong jaws to break through the chest cavity and puncture the heart and lungs of its prey.
The animal faced competition from whale-eating cetaceans, such as Livyatan and other macroraptorial sperm whales and possibly smaller ancestral killer whales. As the shark preferred warmer waters, it is thought that oceanic cooling associated with the onset of the ice ages, coupled with the lowering of sea levels and resulting loss of suitable nursery areas, may have also contributed to its decline. A reduction in the diversity of baleen whales and a shift in their distribution toward polar regions may have reduced megalodon’s primary food source. The shark’s extinction coincides with a gigantism trend in baleen whales.
Taxonomy
Naming
The depiction of a shark’s head by Nicolas Steno in his work The Head of a Shark Dissected
According to Renaissance accounts, gigantic triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolas Steno, who recognized them as shark teeth, and famously produced a depiction of a shark’s head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a megalodon tooth.[16][17][18]
Swiss naturalist Louis Agassiz gave this shark its initial scientific name, Carcharodon megalodon, in his 1843 work Recherches sur les poissons fossiles, based on tooth remains.[1][19] English paleontologist Edward Charlesworth in his 1837 paper used the name Carcharias megalodon, while citing Agassiz as the author, indicating that Agassiz described the species prior to 1843. English paleontologist Charles Davies Sherborn in 1928 listed an 1835 series of articles by Agassiz as the first scientific description of the shark.[20] The specific name megalodon translates to «big tooth», from Ancient Greek: μέγας, romanized: (mégas), lit. ‘big, mighty’ and ὀδούς (odoús), «tooth».[21][22] The teeth of megalodon are morphologically similar to those of the great white shark (Carcharodon carcharias), and on the basis of this observation, Agassiz assigned megalodon to the genus Carcharodon.[19]
There was one apparent description of the shark in 1881 classifying it as Selache manzonii.[23]
Evolution
While the earliest megalodon remains have been reported from the Late Oligocene, around 28 million years ago (Mya),[24][25] there is disagreement as to when it appeared, with dates ranging to as young as 16 mya.[26] It has been thought that megalodon became extinct around the end of the Pliocene, about 2.6 Mya;[26][27] claims of Pleistocene megalodon teeth, younger than 2.6 million years old, are considered unreliable.[27] A 2019 assessment moves the extinction date back to earlier in the Pliocene, 3.6 Mya.[28]
Megalodon is now considered to be a member of the family Otodontidae, genus Otodus, as opposed to its previous classification into Lamnidae, genus Carcharodon.[26][12][27][6][7] Megalodon’s classification into Carcharodon was due to dental similarity with the great white shark, but most authors currently believe that this is due to convergent evolution. In this model, the great white shark is more closely related to the extinct broad-toothed mako (Isurus hastalis) than to megalodon, as evidenced by more similar dentition in those two sharks; megalodon teeth have much finer serrations than great white shark teeth. The great white shark is more closely related to the mako shark (Isurus spp.), with a common ancestor around 4 Mya.[19][29] Proponents of the former model, wherein megalodon and the great white shark are more closely related, argue that the differences between their dentition are minute and obscure.[30]: 23–25
The genus Carcharocles currently contains four species: C. auriculatus, C. angustidens, C. chubutensis, and C. megalodon.[31]: 30–31 The evolution of this lineage is characterized by the increase of serrations, the widening of the crown, the development of a more triangular shape, and the disappearance of the lateral cusps.[31]: 28–31 [32] The evolution in tooth morphology reflects a shift in predation tactics from a tearing-grasping bite to a cutting bite, likely reflecting a shift in prey choice from fish to cetaceans.[33] Lateral cusplets were finally lost in a gradual process that took roughly 12 million years during the transition between C. chubutensis and C. megalodon.[33] The genus was proposed by D. S. Jordan and H. Hannibal in 1923 to contain C. auriculatus. In the 1980s, megalodon was assigned to Carcharocles.[19][31]: 30 Before this, in 1960, the genus Procarcharodon was erected by French ichthyologist Edgard Casier, which included those four sharks and was considered separate from the great white shark. It is now considered a junior synonym of Carcharocles.[31]: 30 The genus Palaeocarcharodon was erected alongside Procarcharodon to represent the beginning of the lineage, and, in the model wherein megalodon and the great white shark are closely related, their last common ancestor. It is believed to be an evolutionary dead-end and unrelated to the Carcharocles sharks by authors who reject that model.[30]: 70
The great white shark (Carcharodon carcharias) and megalodon were previously thought to be close relatives.[19][29]
Another model of the evolution of this genus, also proposed by Casier in 1960, is that the direct ancestor of the Carcharocles is the shark Otodus obliquus, which lived from the Paleocene through the Miocene epochs, 60 to 13 Mya.[29][32] The genus Otodus is ultimately derived from Cretolamna, a shark from the Cretaceous period.[6][34] In this model, O. obliquus evolved into O. aksuaticus, which evolved into C. auriculatus, and then into C. angustidens, and then into C. chubutensis, and then finally into C. megalodon.
Another model of the evolution of Carcharocles, proposed in 2001 by paleontologist Michael Benton, is that the three other species are actually a single species of shark that gradually changed over time between the Paleocene and the Pliocene, making it a chronospecies.[31]: 17 [25][35] Some authors suggest that C. auriculatus, C. angustidens, and C. chubutensis should be classified as a single species in the genus Otodus, leaving C. megalodon the sole member of Carcharocles.[25][36]
The genus Carcharocles may be invalid, and the shark may actually belong in the genus Otodus, making it Otodus megalodon.[4] A 1974 study on Paleogene sharks by Henri Cappetta erected the subgenus Megaselachus, classifying the shark as Otodus (Megaselachus) megalodon, along with O. (M.) chubutensis. A 2006 review of Chondrichthyes elevated Megaselachus to genus, and classified the sharks as Megaselachus megalodon and M. chubutensis.[4] The discovery of fossils assigned to the genus Megalolamna in 2016 led to a re-evaluation of Otodus, which concluded that it is paraphyletic, that is, it consists of a last common ancestor but it does not include all of its descendants. The inclusion of the Carcharocles sharks in Otodus would make it monophyletic, with the sister clade being Megalolamna.[6]
The cladogram below represents the hypothetical relationships between megalodon and other sharks, including the great white shark. Modified from Shimada et al. (2016),[6] Ehret et al., (2009),[29] and the findings of Siversson et al. (2013).[38][39]
Biology
Appearance
One interpretation on how megalodon appeared was that it was a robust-looking shark, and may have had a similar build to the great white shark. The jaws may have been blunter and wider than the great white, and the fins would have also been similar in shape, though thicker due to its size. It may have had a pig-eyed appearance, in that it had small, deep-set eyes.[40]
Another interpretation is that megalodon bore a similarity to the whale shark (Rhincodon typus) or the basking shark (Cetorhinus maximus). The tail fin would have been crescent-shaped, the anal fin and second dorsal fin would have been small, and there would have been a caudal keel present on either side of the tail fin (on the caudal peduncle). This build is common in other large aquatic animals, such as whales, tuna, and other sharks, in order to reduce drag while swimming. The head shape can vary between species as most of the drag-reducing adaptations are toward the tail-end of the animal.[31]: 35–36
Sculpture in the Museum of Evolution in Puebla, Mexico
Since Carcharocles is derived from Otodus, and the two had teeth that bear a close similarity to those of the sand tiger shark (Carcharias taurus), megalodon may have had a build more similar to the sand tiger shark than to other sharks. This is unlikely since the sand tiger shark is a carangiform swimmer which requires faster movement of the tail for propulsion through the water than the great white shark, a thunniform swimmer.[31]: 35–36 [41]
Size
Due to fragmentary remains, there have been many contradictory size estimates for megalodon, as they can only be drawn from fossil teeth and vertebrae.[42]: 87 [43] The great white shark has been the basis of reconstruction and size estimation, as it is regarded as the best analogue to megalodon. Several total length estimation methods have been produced from comparing megalodon teeth and vertebrae to those of the great white.[40][44][10][7]
Size comparison of the great white and whale shark to estimates for megalodon
Proportions of megalodon at lengths of 3 m (10 ft), 8 m (26 ft), and 16 m (52 ft), extrapolated from extant relatives, with a 1.65 m (5 ft 5 in) diver
Megalodon size estimates vary depending on the method used, with maximum total length estimates ranging from 14.2–20.3 meters (47–67 ft).[40][10][7] A 2015 study estimated the average total body length at 10.5 meters (34 ft), calculated from 544 megalodon teeth, found throughout geological time and geography, including adults and juveniles.[11][12] In comparison, large great white sharks are generally around 6 meters (20 ft) in length, with a few contentious reports suggesting larger sizes.[45][46][40] The whale shark is the largest living fish, with one large female reported with a precaudal length of 15 meters (49 ft) and an estimated total length of 18.8 meters (62 ft).[45][47] It is possible that different populations of megalodon around the globe had different body sizes and behaviors due to different ecological pressures.[12] Megalodon is thought to have been the largest macropredatory shark that ever lived.[40]
«A C. megalodon about 16 meters long would have weighed about 48 metric tons (53 tons). A 17-meter (56-foot) C. megalodon would have weighed about 59 metric tons (65 tons), and a 20.3-meter (67 foot) monster would have topped off at 103 metric tons (114 tons).»[13]
In his 2015 book, The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution, Donald Prothero proposed the body mass estimates for different individuals of different length by extrapolating from a vertebral centra based on the dimensions of the great white,[13] a methodology also used for the 2008 study which supports the maximum mass estimate.[14]
In 2020, Cooper and his colleagues reconstructed a 2D model of megalodon based on the dimensions of all the extant lamnid sharks and suggested that a 16 meters (52 ft) long megalodon would have had a 4.65 m (15.3 ft) long head, 1.41 m (4 ft 8 in) tall gill slits, a 1.62 m (5 ft 4 in) tall dorsal fin, 3.08 m (10 ft 1 in) long pectoral fins, and a 3.85 m (12 ft 8 in) tall tail fin.[8] In 2022, Cooper and his colleagues also reconstructed a 3D model with the same basis as the 2020 study, resulting in a body mass estimate of 61.56 metric tons (67.86 short tons) for a 16 meters (52 ft) long megalodon (higher than the previous estimates); a vertebral column specimen named IRSNB P 9893 (formerly IRSNB 3121), belonging to a 46 year old individual from Belgium, was used for extrapolation. An individual of this size would have required 98,175 kcal per day, 20 times more than what the adult great white requires.[15]
Mature male megalodon may have had a body mass of 12.6 to 33.9 metric tons (13.9 to 37.4 short tons), and mature females may have been 27.4 to 59.4 metric tons (30.2 to 65.5 short tons), assuming that males could range in length from 10.5 to 14.3 meters (34 to 47 ft) and females 13.3 to 17 meters (44 to 56 ft).[40]
A 2015 study linking shark size and typical swimming speed estimated that megalodon would have typically swum at 18 kilometers per hour (11 mph)–assuming that its body mass was typically 48 metric tons (53 short tons)–which is consistent with other aquatic creatures of its size, such as the fin whale (Balaenoptera physalus) which typically cruises at speeds of 14.5 to 21.5 km/h (9.0 to 13.4 mph).[48] In 2022, Cooper and his colleagues converted this calculation into relative cruising speed (body lengths per second), resulting in an mean absolute cruising speed of 5 kilometers per hour (3.1 mph) and a mean relative cruising speed of 0.09 body lengths per second for a 16 meters (52 ft) long megalodon; the authors found their mean absolute cruising speed to be faster than any extant lamnid sharks and their mean relative cruising speed to be slower, consistent with previous estiamtes.[15]
Its large size may have been due to climatic factors and the abundance of large prey items, and it may have also been influenced by the evolution of regional endothermy (mesothermy) which would have increased its metabolic rate and swimming speed. The otodontid sharks have been considered to have been ectotherms, so on that basis megalodon would have been ectothermic. However, the largest contemporary ectothermic sharks, such as the whale shark, are filter feeders, while lamnids are now known to be regional endotherms, implying some metabolic correlations with a predatory lifestyle. These considerations, as well as tooth oxygen isotopic data and the need for higher burst swimming speeds in macropredators of endothermic prey than ectothermy would allow, imply that otodontids, including megalodon, were probably regional endotherms.[49]
In 2020, Shimada and colleagues suggested large size was instead due to intrauterine cannibalism, where the larger fetus eats the smaller fetus, resulting in progressively larger and larger fetuses, requiring the mother to attain even greater size as well as caloric requirements which would have promoted endothermy. Males would have needed to keep up with female size in order to still effectively copulate (which probably involved latching onto the female with claspers, like modern cartilaginous fish).[50]
Maximum estimates
The first attempt to reconstruct the jaw of megalodon was made by Bashford Dean in 1909, displayed at the American Museum of Natural History. From the dimensions of this jaw reconstruction, it was hypothesized that megalodon could have approached 30 meters (98 ft) in length. Dean had overestimated the size of the cartilage on both jaws, causing it to be too tall.[51][52]
Tooth compared to hand
In 1973, John E. Randall, an ichthyologist, used the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) to measure the length of the shark, yielding a maximum length of about 13 meters (43 ft).[53] However, tooth enamel height does not necessarily increase in proportion to the animal’s total length.[30]: 99
In 1994, marine biologists Patrick J. Schembri and Stephen Papson opined that O. megalodon may have approached a maximum of around 24 to 25 meters (79 to 82 ft) in total length.[54][55]
In 1996, shark researchers Michael D. Gottfried, Leonard Compagno, and S. Curtis Bowman proposed a linear relationship between the great white shark’s total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in meters = − (0.096) × [UA maximum height (mm)]-(0.22).[56][40] Using this tooth height regression equation, the authors estimated a total length of 15.9 meters (52 ft) based on a tooth 16.8 centimeters (6.6 in) tall, which the authors considered a conservative maximum estimate. They also compared the ratio between the tooth height and total length of large female great whites to the largest megalodon tooth. A 6-meter (20 ft) long female great white, which the authors considered the largest ‘reasonably trustworthy’ total length, produced an estimate of 16.8 meters (55 ft). However, based on the largest female great white reported, at 7.1 meters (23 ft), they estimated a maximum estimate of 20.2 meters (66 ft).[40]
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimeter (0.39 in) of root width, there are approximately 1.4 meters (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah’s possession had a root width of about 12 centimeters (4.7 in), which yielded 16.5 meters (54 ft) in total length.[31]: 88
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length after conducting anatomical analysis of several specimens, allowing any sized tooth to be used. Shimada stated that the previously proposed methods were based on a less-reliable evaluation of the dental homology between megalodon and the great white shark, and that the growth rate between the crown and root is not isometric, which he considered in his model. Using this model, the upper anterior tooth possessed by Gottfried and colleagues corresponded to a total length of 15 meters (49 ft).[57] Among several specimens found in the Gatún Formation of Panama, one upper lateral tooth was used by other researchers to obtain a total length estimate of 17.9 meters (59 ft) using this method.[36][58]
In 2019, Shimada revisited the size of megalodon and discouraged using non-anterior teeth for estimations, noting that the exact position of isolated non-anterior teeth is difficult to identify. Shimada provided maximum total length estimates using the largest anterior teeth available in museums. The tooth with the tallest crown height known to Shimada, NSM PV-19896, produced a total length estimate of 14.2 meters (47 ft). The tooth with the tallest total height, FMNH PF 11306, was reported at 16.8 centimeters (6.6 in). However, Shimada remeasured the tooth and found it actually to measure 16.2 centimeters (6.4 in). Using the total height tooth regression equation proposed by Gottfried and colleagues produced an estimate of 15.3 meters (50 ft).[7][10]
In 2021, Victor J. Perez, Ronny M. Leder, and Teddy Badaut proposed a method of estimating total length of megalodon from the sum of the tooth crown widths. Using more complete megalodon dentitions, they reconstructed the dental formula and then made comparisons to living sharks. The researchers noted that the 2002 Shimada crown height equations produce wildly varying results for different teeth belonging to the same shark (range of error of ± 9 metres (30 ft)), casting doubt on some of the conclusions of previous studies using that method. Using the largest tooth available to the authors, GHC 6, with a crown width of 13.3 centimeters (5.2 in), they estimated a maximum body length of approximately 20 meters (66 ft), with a range of error of approximately ± 3.5 metres (11 ft).[10] This maximum length estimate was also supported by Cooper and his colleagues in 2022.[15]
There are anecdotal reports of teeth larger than those found in museum collections.[7] Gordon Hubbell from Gainesville, Florida, possesses an upper anterior megalodon tooth whose maximum height is 18.4 centimeters (7.25 in), one of the largest known tooth specimens from the shark.[59] In addition, a 2.7-by-3.4-meter (9 by 11 ft) megalodon jaw reconstruction developed by fossil hunter Vito Bertucci contains a tooth whose maximum height is reportedly over 18 centimeters (7 in).[60]
Teeth and bite force
The most common fossils of megalodon are its teeth. Diagnostic characteristics include a triangular shape, robust structure, large size, fine serrations, a lack of lateral denticles, and a visible V-shaped neck (where the root meets the crown).[30]: 55 [36] The tooth met the jaw at a steep angle, similar to the great white shark. The tooth was anchored by connective tissue fibers, and the roughness of the base may have added to mechanical strength.[61] The lingual side of the tooth, the part facing the tongue, was convex; and the labial side, the other side of the tooth, was slightly convex or flat. The anterior teeth were almost perpendicular to the jaw and symmetrical, whereas the posterior teeth were slanted and asymmetrical.[62]
Megalodon teeth can measure over 180 millimeters (7.1 in) in slant height (diagonal length) and are the largest of any known shark species,[31]: 33 implying it was the largest of all macropredatory sharks.[40] In 1989, a nearly complete set of megalodon teeth was discovered in Saitama, Japan. Another nearly complete associated megalodon dentition was excavated from the Yorktown Formations in the United States, and served as the basis of a jaw reconstruction of megalodon at the National Museum of Natural History (USNM). Based on these discoveries, an artificial dental formula was put together for megalodon in 1996.[30]: 55 [63]
The dental formula of megalodon is: 2.1.7.43.0.8.4. As evident from the formula, megalodon had four kinds of teeth in its jaws: anterior, intermediate, lateral, and posterior. Megalodon’s intermediate tooth technically appears to be an upper anterior and is termed as «A3» because it is fairly symmetrical and does not point mesially (side of the tooth toward the midline of the jaws where the left and right jaws meet). Megalodon had a very robust dentition,[30]: 20–21 and had over 250 teeth in its jaws, spanning 5 rows.[31]: iv It is possible that large megalodon individuals had jaws spanning roughly 2 meters (6.6 ft) across.[31]: 129 The teeth were also serrated, which would have improved efficiency in cutting through flesh or bone.[19][31]: 1 The shark may have been able to open its mouth to a 75° angle, though a reconstruction at the USNM approximates a 100° angle.[40]
In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5-meter (8.2 ft) long specimen, and then isometrically scaled the results for its maximum size and the conservative minimum and maximum body mass of megalodon. They placed the bite force of the latter between 108,514 to 182,201 newtons (24,395 to 40,960 lbf) in a posterior bite, compared to the 18,216 newtons (4,095 lbf) bite force for the largest confirmed great white shark, and 7,400 newtons (1,700 lbf) for the placoderm fish Dunkleosteus. In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the force generated, which would probably have caused the total force experienced by prey to be higher than the estimate.[14][64]
In 2021, Antonio Ballell and Humberto Ferrón used Finite Element Analysis modeling to examine the stress distribution of three types of megalodon teeth and closely related mega-toothed species when exposed to anterior and lateral forces, the latter of which would be generated when a shark shakes its head to tear through flesh. The resulting simulations identified higher levels of stress in megalodon teeth under lateral force loads compared to its precursor species such as O. obliquus and O. angusteidens when tooth size was removed as a factor. This suggests that megalodon teeth were of a different functional significance than previously expected, challenging prior interpretations that megalodon’s dental morphology was primarily driven by a dietary shift towards marine mammals. Instead, the authors proposed that it was a byproduct of an increase in body size caused by heterochronic selection.[65]
Internal anatomy
Megalodon is represented in the fossil record by teeth, vertebral centra, and coprolites.[40][66] As with all sharks, the skeleton of megalodon was formed of cartilage rather than bone; consequently most fossil specimens are poorly preserved.[67] To support its large dentition, the jaws of megalodon would have been more massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. Its chondrocranium, the cartilaginous skull, would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size.[40]
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp Basin, Belgium, in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimeters (2.2 in) to 155 millimeters (6 in) in diameter. The shark’s vertebrae may have gotten much bigger, and scrutiny of the specimen revealed that it had a higher vertebral count than specimens of any known shark, possibly over 200 centra; only the great white approached it.[40] Another partially preserved vertebral column of a megalodon was excavated from the Gram Formation in Denmark in 1983, which comprises 20 vertebral centra, with the centra ranging from 100 millimeters (4 in) to 230 millimeters (9 in) in diameter.[61]
The coprolite remains of megalodon are spiral-shaped, indicating that the shark may have had a spiral valve, a corkscrew-shaped portion of the lower intestines, similar to extant lamniform sharks. Miocene coprolite remains were discovered in Beaufort County, South Carolina, with one measuring 14 cm (5.5 in).[66]
Gottfried and colleagues reconstructed the entire skeleton of megalodon, which was later put on display at the Calvert Marine Museum in the United States and the Iziko South African Museum.[40][32] This reconstruction is 11.3 meters (37 ft) long and represents a mature male,[40]: 61 based on the ontogenetic changes a great white shark experiences over the course of its life.[40]: 65
Paleobiology
Range and habitat
Megalodon had a cosmopolitan distribution;[26][58] its fossils have been excavated from many parts of the world, including Europe, Africa, the Americas, and Australia.[30]: 67 [68] It most commonly occurred in subtropical to temperate latitudes.[26][30]: 78 It has been found at latitudes up to 55° N; its inferred tolerated temperature range was 1–24 °C (34–75 °F).[note 1] It arguably had the capacity to endure such low temperatures due to mesothermy, the physiological capability of large sharks to maintain a higher body temperature than the surrounding water by conserving metabolic heat.[26]
Megalodon inhabited a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult megalodon were not abundant in shallow water environments, and mostly inhabited offshore areas. Megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.[31]: 33 [70]
Fossil remains show a trend for specimens to be larger on average in the Southern Hemisphere than in the Northern, with mean lengths of 11.6 and 9.6 meters (38 and 31 ft), respectively; and also larger in the Pacific than the Atlantic, with mean lengths of 10.9 and 9.5 meters (36 and 31 ft) respectively. They do not suggest any trend of changing body size with absolute latitude, or of change in size over time (although the Carcharocles lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 meters (34 ft), with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.[12]
Locations of fossils
Megalodon had a global distribution and fossils of the shark have been found in many places around the world, bordering all oceans of the Neogene.[71]
| Epoch | Formation | State | Continent |
|---|---|---|---|
| Pliocene | Luanda Formation | Africa | |
| Africa | |||
| Africa | |||
| Castell’Arquato Formation | Europe | ||
| Arenas de Huelva Formation | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Touril Complex Formation | Europe | ||
| Red Crag Formation | Europe | ||
| San Mateo Formation | North America | ||
| Towsley Formation | North America | ||
| Bone Valley Formation | North America | ||
| Tamiami Formation | North America | ||
| Yorktown Formation | North America | ||
| Highlands Formation | North America | ||
| Refugio Formation | North America | ||
| San Diego Formation | North America | ||
| Tirabuzon Formation | North America | ||
| Onzole Formation | South America | ||
| Paraguaná Formation | South America | ||
| Black Rock Sandstone | Oceania | ||
| Cameron Inlet Formation | Oceania | ||
| Grange Burn Formation | Oceania | ||
| Loxton Sand Formation | Oceania | ||
| Whaler’s Bluff Formation | Oceania | ||
| Tangahoe Formation | Oceania | ||
| Miocene | |||
| Africa | |||
| Madagascar Basin | Africa | ||
| Africa | |||
| Varswater Formation | Africa | ||
| Baripada Limestone | Asia | ||
| Arakida Formation | Asia | ||
| Bihoku Group | Asia | ||
| Fujina Formation | Asia | ||
| Hannoura Formation | Asia | ||
| Hongo Formation | Asia | ||
| Horimatsu Formation | Asia | ||
| Ichishi Formation | Asia | ||
| Kurahara Formation | Asia | ||
| Maenami Formation | Asia | ||
| Matsuyama Group | Asia | ||
| Sekinobana Formation | Asia | ||
| Suso Formation | Asia | ||
| Takakubo Formation | Asia | ||
| Tonokita Formation | Asia | ||
| Tsurushi Formation | Asia | ||
| Wajimazaki Formation | Asia | ||
| Yoshii Formation | Asia | ||
| Asia | |||
| Burgeschleinitz Formation | Europe | ||
| Melker Sand Formation | Europe | ||
| Rzehakia Formation | Europe | ||
| Weissenegg Formation | Europe | ||
| Antwerpen Sands Member | Europe | ||
| Europe | |||
| Hrušky Formation | Europe | ||
| Gram Formation | Europe | ||
| Aquitaine Basin | Europe | ||
| Europe | |||
| Libano Sandstone | Europe | ||
| Blue Clay Formation | Europe | ||
| Globigerina Limestone[72] | Europe | ||
| Aalten Member | Europe | ||
| Breda Formation | Europe | ||
| Korytnica Clays | Europe | ||
| Leitha Limestone | Europe | ||
| Esbarrondadoiro Formation | Europe | ||
| Filakovo Formation | Europe | ||
| Arjona Formation | Europe | ||
| Calcarenites of Sant Elm | Europe | ||
| Europe | |||
| Monterey Formation | North America | ||
| Puente Formation | North America | ||
| Purisima Formation | North America | ||
| San Mateo Formation | North America | ||
| Santa Margarita Formation | North America | ||
| Temblor Formation | North America | ||
| Topanga Formation | North America | ||
| Bone Valley Formation | North America | ||
| Calvert Formation | North America | ||
| Kirkwood Formation | North America | ||
| North America | |||
| Cojímar Formation | North America | ||
| Kendance Formation | North America | ||
| North America | |||
| Aymamón Limestone | North America | ||
| Almejas Formation | North America | ||
| Carrillo Puerto Formation | North America | ||
| Chagres Formation | North America | ||
| Chucunaque Formation | North America | ||
| Gatún Formation | North America | ||
| Paraná Formation | South America | ||
| Bahía Inglesa Formation | South America | ||
| Castilletes Formation | South America | ||
| Miramar Formation | South America | ||
| Pisco Formation | South America | ||
| Camacho Formation | South America | ||
| Cantaure Formation | South America | ||
| Caujarao Formation | South America | ||
| Socorro Formation | South America | ||
| Urumaco Formation | South America | ||
| Batesford Limestone | Oceania | ||
| Black Rock Sandstone | Oceania | ||
| Gippsland Limestone | Oceania | ||
| Mannum Formation | Oceania | ||
| Morgan Limestone | Oceania | ||
| Port Campbell Limestone | Oceania | ||
| Oceania | |||
| Oceania | |||
| Duho Formation[73] | Asia | ||
| Seogwipo Formation[74] | Asia |
Locations of megalodon fossil discoveries, yellow from the Pliocene and blue from the Miocene[26][71]
Prey relationships
Vertebra of a whale bitten in half by a megalodon with visible gashes from teeth.
Though sharks are generally opportunistic feeders, megalodon’s great size, high-speed swimming capability, and powerful jaws, coupled with an impressive feeding apparatus, made it an apex predator capable of consuming a broad spectrum of animals. Otodus megalodon was probably one of the most powerful predators to have existed.[15] A study focusing on calcium isotopes of extinct and extant elasmobranch sharks and rays revealed that megalodon fed at a higher trophic level than the contemporaneous great white shark («higher up» in the food chain.)[75]
Fossil evidence indicates that megalodon preyed upon many cetacean species, such as dolphins, small whales, cetotheres, squalodontids (shark toothed dolphins), sperm whales, bowhead whales, and rorquals.[51][76][77] In addition to this, they also targeted seals, sirenians, and sea turtles.[70] The shark was an opportunist and piscivorous, and it would have also gone after smaller fish and other sharks.[51] Many whale bones have been found with deep gashes most likely made by their teeth.[30]: 75 Various excavations have revealed megalodon teeth lying close to the chewed remains of whales,[30]: 75 [32] and sometimes in direct association with them.[78]
The feeding ecology of megalodon appears to have varied with age and between sites, like the modern great white shark. It is plausible that the adult megalodon population off the coast of Peru targeted primarily cetothere whales 2.5 to 7 meters (8.2 to 23 ft) in length and other prey smaller than itself, rather than large whales in the same size class as themselves.[76] Meanwhile, juveniles likely had a diet that consisted more of fish.[36][79]
Competition
Megalodon faced a highly competitive environment.[80] Its position at the top of the food chain[81] probably had a significant impact on the structuring of marine communities.[80][82] Fossil evidence indicates a correlation between megalodon and the emergence and diversification of cetaceans and other marine mammals.[30]: 78 [80] Juvenile megalodon preferred habitats where small cetaceans were abundant, and adult megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.[30]: 74–75
Megalodon were contemporaneous with whale-eating toothed whales (particularly macroraptorial sperm whales and squalodontidae), which were also probably among the era’s apex predators, and provided competition.[80] Some attained gigantic sizes, such as Livyatan, estimated between 13.5 to 17.5 meters (44 to 57 ft). Fossilized teeth of an undetermined species of such physeteroids from Lee Creek Mine, North Carolina, indicate it had a maximum body length of 8–10 m and a maximum lifespan of about 25 years. This is very different from similarly sized modern killer whales that live to 65 years, suggesting that unlike the latter, which are apex predators, these physeteroids were subject to predation from larger species such as megalodon or Livyatan.[83] By the Late Miocene, around 11 Mya, macroraptorials experienced a significant decline in abundance and diversity. Other species may have filled this niche in the Pliocene,[80][84] such as the fossil killer whale Orcinus citoniensis which may have been a pack predator and targeted prey larger than itself,[32][85][86][87] but this inference is disputed,[28] and it was probably a generalist predator rather than a marine mammal specialist.[88]
Megalodon may have subjected contemporaneous white sharks to competitive exclusion, as the fossil records indicate that other shark species avoided regions it inhabited by mainly keeping to the colder waters of the time.[89][30]: 77 In areas where their ranges seemed to have overlapped, such as in Pliocene Baja California, it is possible that megalodon and the great white shark occupied the area at different times of the year while following different migratory prey.[30]: 77 [90] Megalodon probably also had a tendency for cannibalism, much like contemporary sharks.[91]
Feeding strategies
Sharks often employ complex hunting strategies to engage large prey animals. Great white shark hunting strategies may be similar to how megalodon hunted its large prey.[92] Megalodon bite marks on whale fossils suggest that it employed different hunting strategies against large prey than the great white shark.[51]
One particular specimen–the remains of a 9-meter (30 ft) long undescribed Miocene baleen whale–provided the first opportunity to quantitatively analyze its attack behavior. Unlike great whites which target the underbelly of their prey, megalodon probably targeted the heart and lungs, with their thick teeth adapted for biting through tough bone, as indicated by bite marks inflicted to the rib cage and other tough bony areas on whale remains.[51] Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans, for example cetotheres, suggest that they were rammed with great force from below before being killed and eaten, based on compression fractures.[92]
There is also evidence that a possible separate hunting strategy existed for attacking raptorial sperm whales; a tooth belonging to an undetermined 4 m (13 ft) physeteroid closely resembling those of Acrophyseter discovered in the Nutrien Aurora Phosphate Mine in North Carolina suggests that a megalodon or O. chubutensis may have aimed for the head of the sperm whale in order to inflict a fatal bite, the resulting attack leaving distinctive bite marks on the tooth. While scavenging behavior cannot be ruled out as a possibility, the placement of the bite marks is more consistent with predatory attacks than feeding by scavenging, as the jaw is not a particularly nutritious area to for a shark feed or focus on. The fact that the bite marks were found on the tooth’s roots further suggest that the shark broke the whale’s jaw during the bite, suggesting the bite was extremely powerful. The fossil is also notable as it stands as the first known instance of an antagonistic interaction between a sperm whale and an otodontid shark recorded in the fossil record.[93]
During the Pliocene, larger cetaceans appeared.[94] Megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones and tail vertebrae of large whales from the Pliocene have been found with megalodon bite marks, which suggests that megalodon would immobilize a large whale before killing and feeding on it.[14][51]
Growth and reproduction
In 2010, Ehret estimated that megalodon had a fast growth rate nearly two times that of the extant great white shark. He also estimated that the slowing or cessation of somatic growth in megalodon occurred around 25 years of age, suggesting that this species had an extremely delayed sexual maturity.[95] In 2021, Shimada and colleagues calculated the growth rate of an approximately 9.2 m (30 ft) individual based on the Belgian vertebrate column specimen that presumably contains annual growth rings on three of its vertebrae. They estimated the individual died at 46 years of age, with a growth rate of 16 cm (6.3 in) per year, and a length of 2 m (6 ft 7 in) at birth. For a 15 m (49 ft) individual—which they considered to have been the maximum size attainable—this would equate to a lifespan of 88 to 100 years.[96] However, Cooper and his colleagues in 2022 estimated the length of this 46 year old individual at nearly 16 m (52 ft) based on the 3D reconstruction which resulted in the complete vertebral column to be 11.1 m (36 ft) long; the researchers claimed that this size estimate difference occurred due to the fact that Shimada and his colleagues extrapolated its size only based on the vertebral centra.[15]
Megalodon, like contemporaneous sharks, made use of nursery areas to birth their young in, specifically warm-water coastal environments with large amounts of food and protection from predators.[36] Nursery sites were identified in the Gatún Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands,[97] and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of megalodon also.[98] Infant megalodons were around 3.5 meters (11 ft) at their smallest,[40]: 61 and the pups were vulnerable to predation by other shark species, such as the great hammerhead shark (Sphyrna mokarran) and the snaggletooth shark (Hemipristis serra).[36] Their dietary preferences display an ontogenetic shift:[40]: 65 Young megalodon commonly preyed on fish,[36] sea turtles,[70] dugongs,[31]: 129 and small cetaceans; mature megalodon moved to off-shore areas and consumed large cetaceans.[30]: 74–75
An exceptional case in the fossil record suggests that juvenile megalodon may have occasionally attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-to-7-meter (13 to 23 ft) long Pliocene shark were found on a rib from an ancestral blue or humpback whale that showed evidence of subsequent healing, which is suspected to have been inflicted by a juvenile megalodon.[99][100]
Extinction
Climate change
The Earth experienced a number of changes during the time period megalodon existed which affected marine life. A cooling trend starting in the Oligocene 35 Mya ultimately led to glaciation at the poles. Geological events changed currents and precipitation; among these were the closure of the Central American Seaway and changes in the Tethys Ocean, contributing to the cooling of the oceans. The stalling of the Gulf Stream prevented nutrient-rich water from reaching major marine ecosystems, which may have negatively affected its food sources. The largest fluctuation of sea levels in the Cenozoic era occurred in the Plio-Pleistocene, between around 5 million to 12 thousand years ago, due to the expansion of glaciers at the poles, which negatively impacted coastal environments, and may have contributed to its extinction along with those of several other marine megafaunal species.[101] These oceanographic changes, in particular the sea level drops, may have restricted many of the suitable shallow warm-water nursery sites for megalodon, hindering reproduction.[102] Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.[103][36]
As its range did not apparently extend into colder waters, megalodon may not have been able to retain a significant amount of metabolic heat, so its range was restricted to shrinking warmer waters.[102][77][104] Fossil evidence confirms the absence of megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[30]: 77 However, an analysis of the distribution of megalodon over time suggests that temperature change did not play a direct role in its extinction. Its distribution during the Miocene and Pliocene did not correlate with warming and cooling trends; while abundance and distribution declined during the Pliocene, megalodon did show a capacity to inhabit colder latitudes. It was found in locations with a mean temperature ranging from 12 to 27 °C (54 to 81 °F), with a total range of 1 to 33 °C (34 to 91 °F), indicating that the global extent of suitable habitat should not have been greatly affected by the temperature changes that occurred.[26] This is consistent with evidence that it was a mesotherm.[49]
Changing ecosystem
Marine mammals attained their greatest diversity during the Miocene,[30]: 71 such as with baleen whales with over 20 recognized Miocene genera in comparison to only six extant genera.[106] Such diversity presented an ideal setting to support a super-predator such as megalodon.[30]: 75 By the end of the Miocene, many species of mysticetes had gone extinct;[80] surviving species may have been faster swimmers and thus more elusive prey.[31]: 46 Furthermore, after the closure of the Central American Seaway, tropical whales decreased in diversity and abundance.[104] The extinction of megalodon correlates with the decline of many small mysticete lineages, and it is possible that it was quite dependent on them as a food source.[76] Additionally, a marine megafauna extinction during the Pliocene was discovered to have eliminated 36% of all large marine species including 55% of marine mammals, 35% of seabirds, 9% of sharks, and 43% of sea turtles. The extinction was selective for endotherms and mesotherms relative to poikilotherms, implying causation by a decreased food supply[101] and thus consistent with megalodon being mesothermic.[49] Megalodon may have been too large to sustain itself on the declining marine food resources.[102] The cooling of the oceans during the Pliocene might have restricted the access of megalodon to the polar regions, depriving it of the large whales which had migrated there.[104]
Competition from large odontocetes, such as macropredatory sperm whales which appeared in the Miocene, and a member of genus Orcinus (i.e., Orcinus citoniensis) in the Pliocene,[80][84] is assumed to have contributed to the decline and extinction of megalodon.[26][31]: 46–47 [102][107] But this assumption is disputed:[28] The Orcininae emerged in Mid-Pliocene with O. citoniensis reported from the
Pliocene of Italy,[84][108] and similar forms reported from the Pliocene of England and South Africa,[84] indicating the capacity of these dolphins to cope with increasingly prevalent cold water temperatures in high latitudes.[84] These dolphins were assumed to have been macrophagous in some studies,[26] but on closer inspection, these dolphins are not found to be macrophagous and fed on small fishes instead.[108] On the other hand, gigantic macropredatory sperm whales such as Livyatan-like forms are last reported from Australia and South Africa circa 5 million years ago.[109][110][111] Others, such as Hoplocetus and Scaldicetus also occupied a niche similar to that of modern killer whales but the last of these forms disappeared during the Pliocene.[112][108] Members of genus Orcinus became large and macrophagous in the Pleistocene.[108]
Paleontologist Robert Boessenecker and his colleagues rechecked the fossil record of megalodon for carbon dating errors and concluded that it disappeared circa 3.5 million years ago.[28] Boessenecker and his colleagues further suggest that megalodon suffered range fragmentation due to climatic shifts,[28] and competition with white sharks might have contributed to its decline and extinction.[28] Competition with white sharks is assumed to be a factor in other studies as well,[113][26][107] but this hypothesis warrants further testing.[114] Multiple compounding environmental and ecological factors including climate change and thermal limitations, collapse of prey populations and resource competition with white sharks are believed to have contributed to decline and extinction of megalodon for now.[107]
The extinction of megalodon set the stage for further changes in marine communities. The average body size of baleen whales increased significantly after its disappearance, although possibly due to other, climate-related, causes.[115] Conversely the increase in baleen whale size may have contributed to the extinction of megalodon, as they may have preferred to go after smaller whales; bite marks on large whale species may have come from scavenging sharks. Megalodon may have simply become coextinct with smaller whale species, such as Piscobalaena nana.[105] The extinction of megalodon had a positive impact on other apex predators of the time, such as the great white shark, in some cases spreading to regions where megalodon became absent.[26][113][116]
In popular culture
HMS Challenger discovered megalodon teeth which were erroneously dated to be around 11,000 to 24,000 years old.
Megalodon has been portrayed in many works of fiction, including films and novels, and continues to be a popular subject for fiction involving sea monsters.[117] Reports of supposedly fresh megalodon teeth, such as those found by HMS Challenger in 1873 which were dated in 1959 by the zoologist Wladimir Tschernezky to be around 11,000 to 24,000 years old, helped popularise claims of recent megalodon survival amongst cryptozoologists.[118] These claims are now discredited, and are probably teeth that were well-preserved by a thick mineral-crust precipitate of manganese dioxide, and so had a lower decomposition rate and retained a white color during fossilization. Fossil megalodon teeth can vary in color from off-white to dark browns and greys, and some fossil teeth may have been redeposited into a younger stratum. The claims that megalodon could remain elusive in the depths, similar to the megamouth shark which was discovered in 1976, are unlikely as the shark lived in warm coastal waters and probably could not survive in the cold and nutrient-poor deep sea environment.[119][120]
Contemporary fiction about megalodon surviving into modern times was pioneered by the 1997 novel Meg: A Novel of Deep Terror by Steve Alten and its subsequent sequels. Megalodon subsequently began to feature in films, such as the 2003 direct to video Shark Attack 3: Megalodon, and later The Meg, a 2018 film based on the 1997 book which grossed over $500 million at the box office.[118][121]
Animal Planet’s pseudo-documentary Mermaids: The Body Found included an encounter 1.6 mya between a pod of mermaids and a megalodon.[122] Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives,[123] a controversial docufiction about the creature that presented alleged evidence in order to suggest that megalodons still lived. This program received criticism for being completely fictional and for inadequately disclosing its fictional nature; for example, all of the supposed scientists depicted were paid actors, and there was no disclosure in the documentary itself that it was fictional. In a poll by Discovery, 73% of the viewers of the documentary thought that megalodon was not extinct. In 2014, Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.[51][124][125][126] Despite the criticism from scientists, Megalodon: The Monster Shark Lives was a huge ratings success, gaining 4.8 million viewers, the most for any Shark Week episode up to that point.[127]
Megalodon teeth are the state fossil of North Carolina.[128]
See also
- List of prehistoric cartilaginous fish
- Prehistoric fish
- Largest prehistoric organisms
Notes
- ^ Carbonated bioapatite from a megalodon tooth (of unknown source location) dated to 5.75 ± 0.9 Ma in age has been analyzed for isotope ratios of oxygen (18O/16O) and carbon (13C/12C), using a carbonate clumped-isotope thermometer methodology to yield an estimate of the ambient temperature in that individual’s environment of 19 ± 4 °C.[69]
References
- ^ a b Agassiz, Louis (1843). Recherches sur les poissons fossiles [Research on the fossil fishes] (in French). Neuchatel: Petitpierre. p. 41.
- ^ «Otodus (Megaselachus) megalodon (Agassiz, 1837)». SharkReferences.com. Retrieved 24 October 2017.
- ^ Eastman, C. R. (1904). Maryland Geological Survey. Vol. 2. Baltimore, Maryland: Johns Hopkins University. p. 82.
- ^ a b c Cappetta, H. (1987). «Mesozoic and Cenozoic Elasmobranchii». Handbook of Paleoichthyology. Vol. 3B. München, Germany: Friedrich Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- ^ Hay, O. P. (1901). «Bibliography and Catalogue of the Fossil Vertebrata of North America». Bulletin of the United States Geological Society (179): 308.
- ^ a b c d e f Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (2016). «A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade». Historical Biology. 29 (5): 1–11. doi:10.1080/08912963.2016.1236795. S2CID 89080495.
- ^ a b c d e f Shimada, Kenshu (2019). «The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited». Historical Biology. 33 (7): 1–8. doi:10.1080/08912963.2019.1666840. ISSN 0891-2963. S2CID 208570844.
- ^ a b Cooper, J. A.; Pimiento, C.; Ferrón, H. G.; Benton, M. J. (2020). «Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction». Scientific Reports. 10 (14596): 14596. Bibcode:2020NatSR..1014596C. doi:10.1038/s41598-020-71387-y. PMC 7471939. PMID 32883981.
- ^ «Giant ‘megalodon’ shark extinct earlier than previously thought». Science Daily. 13 February 2019.
- ^ a b c d e Perez, Victor; Leder, Ronny; Badaut, Teddy (2021). «Body length estimation of Neogene macrophagous lamniform sharks (Carcharodon and Otodus) derived from associated fossil dentitions». Palaeontologia Electronica. 24 (1): 1–28. doi:10.26879/1140.
- ^ a b Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c d e Pimiento, C.; Balk, M. A. (2015). «Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators». Paleobiology. 41 (3): 479–490. doi:10.1017/pab.2015.16. PMC 4541548. PMID 26321775.
- ^ a b c Prothero, Donald R. (25 August 2015). «09. Mega-Jaws: The Largest Fish. Carcharocles». The Story of Life in 25 Fossils: Tales of Intrepid Fossil Hunters and the Wonders of Evolution. New York Chichester, West Sussex: Columbia University Press. pp. 96–110. doi:10.7312/prot17190-010. ISBN 978-0-231-53942-5.
- ^ a b c d e Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?» (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ^ a b c d e f Cooper, J.A.; Hutchinson, J.R.; Bernvi, D.C.; Cliff, G.; Wilson, R.P.; Dicken, M.L.; Menzel, J.; Wroe, S.; Pirlo, J.; Pimiento, C. (2022). «The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling». Science Advances. 8 (33): eabm9424. Bibcode:2022SciA….8M9424C. doi:10.1126/sciadv.abm9424. ISSN 2375-2548. PMC 9385135. PMID 35977007.
- ^ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Westport, Connecticut: Libraries Unlimited. pp. 25–26. ISBN 978-1-59158-265-6. OCLC 230807846.
- ^ Hsu, Kuang-Tai (2009). «The Path to Steno’s Synthesis on the Animal Origin of Glossopetrae». In Rosenburg, G. D. (ed.). The Revolution in Geology from the Renaissance to the Enlightenment. Vol. 203. Boulder, Colorado: Geological Society of America. ISBN 978-0-8137-1203-1. OCLC 608657795.
- ^ Eilperin, J. (2012). Demon Fish. Pantheon Books. p. 43. ISBN 978-0-7156-4352-5.
- ^ a b c d e f Nyberg, K. G.; Ciampaglio C. N.; Wray G. A. (2006). «Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth». Journal of Vertebrate Paleontology. 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. S2CID 53640614.
- ^ Keyes, I. W. (2012). «New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record». New Zealand Journal of Geology and Geophysics. 15 (2): 229. doi:10.1080/00288306.1972.10421956.
- ^ μέγας. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ ὀδούς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ^ Lawley, R. (1881). «Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)» [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (in Italian). 5: 167–172.
- ^ Yabe, H.; Goto, M.; Kaneko, N. (2004). «Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records». The Palaeontological Society of Japan. 75: 7–15.
- ^ a b c Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ a b c d e f g h i j k l Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). «Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms». Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754. S2CID 55776834.
- ^ a b c Pimiento, C.; Clements, C. F. (2014). «When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record». PLOS ONE. 9 (10): e111086. Bibcode:2014PLoSO…9k1086P. doi:10.1371/journal.pone.0111086. PMC 4206505. PMID 25338197.
- ^ a b c d e f Boessenecker, R. W.; Ehret, D. J.; Long, D. J.; Churchill, M.; Martin, E.; Boessenecker, S. J. (2019). «The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific». PeerJ. 7: e6088. doi:10.7717/peerj.6088. PMC 6377595. PMID 30783558.
- ^ a b c d Ehret D. J.; Hubbell G.; Macfadden B. J. (2009). «Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru». Journal of Vertebrate Paleontology. 29 (1): 1–13. doi:10.1671/039.029.0113. JSTOR 20491064. S2CID 129585445.
- ^ a b c d e f g h i j k l m n o p q r Klimley, Peter; Ainley, David (1996). «Evolution». Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. ISBN 978-0-12-415031-7. OCLC 212425118.
- ^ a b c d e f g h i j k l m n o p Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ a b c d e Andres, Lutz (2002). «C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles«. fossilguy.com. Retrieved 16 January 2008.
- ^ a b Perez, V. J.; Godfrey, S. J.; Kent, B. W.; Weems, R. E.; Nance, J. R. (2019). «The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time». Journal of Vertebrate Paleontology. 38 (6): e1546732. doi:10.1080/02724634.2018.1546732.
- ^ Siverson, M.; Lindgren, J.; Newbrey, M.G.; Cederström, P.; Cook, T.D. (2013). «Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type» (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. S2CID 58906204. Archived from the original (PDF) on 19 October 2013.
- ^ Benton, M. J.; Pearson, P. N. (2001). «Speciation in the fossil record». Trends in Ecology and Evolution. 16 (7): 405–411. doi:10.1016/s0169-5347(01)02149-8. PMID 11403874.
- ^ a b c d e f g h Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Vivian, G. (2013). «Research debunks Great White lineage».
- ^ «Palaeontological detective work unravels evolution of megatooth sharks». 2013.
- ^ a b c d e f g h i j k l m n o p q r Gottfried, MD; Compagno, LJV; Bowman, SC (1996). «Size and skeletal anatomy of the giant megatooth shark Carcharodon megalodon«. In Klimley; Ainley (eds.). Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. pp. 55–89. ISBN 978-0124150317.
- ^ «Could Megalodon Have Looked Like a BIG Sandtiger Shark?». Biology of Sharks and Rays. Retrieved 2 September 2017.
- ^ Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- ^ Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). «Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles» (PDF). Caribbean Journal of Science. 44 (3): 279–286. doi:10.18475/cjos.v44i3.a2. S2CID 87154947. Archived from the original (PDF) on 20 July 2011.
- ^ Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna (ed.). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO…510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ a b McClain, Craig R.; Balk, Meghan A.; Benfield, Mark C.; Branch, Trevor A.; Chen, Catherine; Cosgrove, James; Dove, Alistair D.M.; Gaskins, Lindsay C.; Helm, Rebecca R.; Hochberg, Frederick G.; Lee, Frank B. (13 January 2015). «Sizing ocean giants: patterns of intraspecific size variation in marine megafauna». PeerJ. 3: e715. doi:10.7717/peerj.715. ISSN 2167-8359. PMC 4304853. PMID 25649000.
- ^ I., Castro, Jose (2011). Sharks of North America. Oxford University Press, USA. ISBN 978-0-19-978097-6. OCLC 958576172.
- ^ Borrell, Asunción; Aguilar, Alex; Gazo, Manel; Kumarran, R. P.; Cardona, Luis (1 December 2011). «Stable isotope profiles in whale shark (Rhincodon typus) suggest segregation and dissimilarities in the diet depending on sex and size». Environmental Biology of Fishes. 92 (4): 559–567. doi:10.1007/s10641-011-9879-y. ISSN 1573-5133. S2CID 37683420.
- ^ Jacoby, D. M. P.; Siriwat, P.; Freeman, R.; Carbone, C. (2015). «Is the scaling of swim speed in sharks driven by metabolism?». Biology Letters. 12 (10): 20150781. doi:10.1098/rsbl.2015.0781. PMC 4707698. PMID 26631246.
- ^ a b c Ferrón, H. G. (2017). «Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks». PLOS ONE. 12 (9): e0185185. Bibcode:2017PLoSO..1285185F. doi:10.1371/journal.pone.0185185. PMC 5609766. PMID 28938002.
- ^ Shimada, K.; Becker, M. A.; Griffiths, M. L. (2020). «Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to ‘off-the-scale’ gigantism of the megatooth shark, Otodus megalodon«. Historical Biology. 33 (11): 1–17. doi:10.1080/08912963.2020.1812598.
- ^ a b c d e f g Prothero, D. R. (2015). «Mega-Jaws». The Story of Life in 25 Fossils. New York, New York: Columbia University Press. pp. 96–110. ISBN 978-0-231-17190-8. OCLC 897505111.
- ^ Helfman, G.; Burgess, G. H. (2014). Sharks: The Animal Answer Guide. Baltimore, Maryland: Johns Hopkins University Press. p. 19. ISBN 978-1-4214-1310-5. OCLC 903293986.
- ^ Randall, John E. (1973). «Size of the Great White Shark (Carcharodon)». Science Magazine. 181 (4095): 169–170. Bibcode:1973Sci…181..169R. doi:10.1126/science.181.4095.169. PMID 17746627. S2CID 36607712.
- ^ Schembri, Patrick (1994). «Malta’s Natural Heritage». Natural Heritage. In: 105–124.
- ^ Papson, Stephen (1992). «Copyright: Cross the Fin Line of Terror». Journal of American Culture. 15 (4): 67–81. doi:10.1111/j.1542-734X.1992.1504_67.x.
- ^ Gottfried, M. D.; Fordyce, R. E. (2001). «An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships». Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. S2CID 86092645. Archived from the original on 17 December 2019. Retrieved 6 November 2017.
- ^ Shimada, Kenshu (2002). «The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)». Journal of Fossil Research. 35 (2): 28–33.
- ^ a b Pimiento, Catalina; Gerardo González-Barba; Dana J. Ehret; Austin J. W. Hendy; Bruce J. MacFadden; Carlos Jaramillo (2013). «Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama» (PDF). Journal of Paleontology. 87 (5): 755–774. doi:10.1666/12-117. S2CID 45662900. Archived from the original (PDF) on 29 October 2013.
- ^ Crane, B. (2017). «A Prehistoric Killer, Buried in Muck». The New Yorker. Retrieved 10 December 2017.
- ^ Mustain, A. (2011). «For Sale: World’s Largest Shark Jaws». LiveScience. Retrieved 31 August 2017.
- ^ a b Almgreen, S. E. Bendix (15 November 1983). «Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn». Bulletin of the Geological Society of Denmark. 32: 1–32. CiteSeerX 10.1.1.514.1782. doi:10.37570/bgsd-1983-32-01. S2CID 53311833. NAID 10012345550.
- ^ Reolid, M.; Molina, J. M. (2015). «Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain)». Estudios Geológicos. 71 (2): e032. doi:10.3989/egeol.41828.342.
- ^ Uyeno, T.; Sakamoto, O.; Sekine, H. (1989). «The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan». Saitama Museum of Natural History Bulletin. 7: 73–85.
- ^ Anderson, P.S.L.; Westneat, M. (2009). «A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)». Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1. S2CID 86203770.
- ^ Ballell, A.; Ferrón, H.G. (2021). «Biomechanical insights into the dentition of megatooth sharks (Lamniformes: Otodontidae)». Scientific Reports. 11 (1232): 1232. doi:10.1038/s41598-020-80323-z. PMC 7806677. PMID 33441828.
- ^ a b Stringer, G. L.; King, L. (2012). «Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana». New Mexico Museum of Natural History and Science, Bulletin. Vertebrate Corpolites. 57: 301.
- ^ «Megalodon Shark Facts and Information: The Details». fossilguy.com. Retrieved 18 September 2017.
- ^ Fitzgerald, Erich (2004). «A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia». Memoirs of Museum Victoria. 61 (2): 183–208. doi:10.24199/j.mmv.2004.61.12.
- ^ Löffler, N.; Fiebig, J.; Mulch, A.; Tütken, T.; Schmidt, B.C.; Bajnai, D.; Conrad, A.C.; Wacker, U.; Böttcher, M.E. (2019). «Refining the temperature dependence of the oxygen and clumped isotopic compositions of structurally bound carbonate in apatite». Geochimica et Cosmochimica Acta. 253: 19–38. Bibcode:2019GeCoA.253…19L. doi:10.1016/j.gca.2019.03.002. S2CID 107992832.
- ^ a b c Aguilera O.; Augilera E. R. D. (2004). «Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage». Caribbean Journal of Science. 40 (3): 362–368.
- ^ a b «Carcharocles megalodon«. Fossilworks. Retrieved 28 August 2017 from the Paleobiology Database.
{{cite web}}: CS1 maint: postscript (link) - ^ «Young Prince George gifted 23m-year-old tooth from extinct shark found in Malta». Times of Malta. 27 September 2020. Retrieved 28 September 2020.
- ^ Yun, C. (2020). «New example of Cosmopolitodus hastalis (Lamniformes, Lamnidae) from the Miocene South Korea». Zoodiversity. 54 (5): 433–438. doi:10.15407/zoo2020.05.433. S2CID 229274996.
- ^ Choi, Seung; Lee, Yuong-Nam (2017). «A review of vertebrate body fossils from the Korean Peninsula and perspectives». Geosciences Journal. 21 (6): 867–889. Bibcode:2017GescJ..21..867C. doi:10.1007/s12303-017-0040-6. ISSN 1226-4806. S2CID 133835817.
- ^ Martin, J. E.; Tacail, T.; Sylvain, A.; Catherine, G.; Vincent, B. (2015). «Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs». Chemical Geology. 415: 118–125. Bibcode:2015ChGeo.415..118M. doi:10.1016/j.chemgeo.2015.09.011.
- ^ a b c Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ a b Morgan, Gary S. (1994). «Whither the giant white shark?» (PDF). Paleontology Topics. 2 (3): 1–2. Archived from the original (PDF) on 22 July 2016.
- ^ Augilera, Orangel A.; García, Luis; Cozzuol, Mario A. (2008). «Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation». Paläontologische Zeitschrift. 82 (2): 204–208. doi:10.1007/BF02988410. ISSN 0038-2353. S2CID 84251638.
- ^ Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). «The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)». Journal of South American Earth Sciences. 73: 168–190. Bibcode:2017JSAES..73..168L. doi:10.1016/j.jsames.2016.12.010.
- ^ a b c d e f g h Lambert, O.; Bianucci, G.; Post, P.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. (2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020. S2CID 4369352.
- ^ Compagno, Leonard J. V. (1989). «Alternative life-history styles of cartilaginous fishes in time and space». Environmental Biology of Fishes. 28 (1–4): 33–75. doi:10.1007/BF00751027. S2CID 22527888.
- ^ Ferretti, Francesco; Boris Worm; Gregory L. Britten; Michael R. Heithaus; Heike K. Lotze1 (2010). «Patterns and ecosystem consequences of shark declines in the ocean» (PDF). Ecology Letters. 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. Archived from the original (PDF) on 6 July 2011. Retrieved 19 February 2011.
- ^ Gilbert, K.N.; Ivany, L.C.; Uhen, M.D. (2018). «Living fast and dying young: life history and ecology of a Neogene sperm whale». Journal of Vertebrate Paleontology. 38 (2): e1439038. doi:10.1080/02724634.2018.1439038. S2CID 89750852.
- ^ a b c d e Heyning, John; Dahlheim, Marilyn (1988). «Orcinus orca» (PDF). Mammalian Species (304): 1–9. doi:10.2307/3504225. JSTOR 3504225. S2CID 253914153. Archived from the original (PDF) on 5 December 2010.
- ^ Bianucci, Giovanni; Walter, Landini (2006). «Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy». Zoological Journal of the Linnean Society. 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x.
- ^ Lindberg, D. R.; Pyenson, N. D. (2006). «Evolutionary Patterns in Cetacea: Fishing Up Prey Size through Deep Time». Whales, Whaling, and Ocean Ecosystems. University of California Press. p. 77. ISBN 978-0-520-24884-7.
- ^ Boessenecker, R. W. (2013). «A new marine vertebrate assemblage from the Late Neogene Purisima Formation in Central California, part II: Pinnipeds and Cetaceans». Geodiversitas. 35 (4): 815–940. doi:10.5252/g2013n4a5. S2CID 85940452.
- ^ Bianucci, G. (1997). «Hemisyntrachelus cortesii (Cetacea, Delphinidae) from the Pliocene Sediments of Campore Quarry (Salsomaggiori Terme, Italy». Bollettino della Societa Paleontologica Italiana. 36 (1): 75–83).
- ^ Antunes, M.T.; Legoinha, P.; Balbing, A. (2015). «Megalodon, mako shark and planktonic foraminifera from the continental shelf off Portugal and their age». Geologica Acta. 13: 181–190.
- ^ «Paleoecology of Megalodon and the White Shark». Biology of Sharks and Rays. Retrieved 1 October 2017.
- ^ Tanke, Darren; Currie, Philip (1998). «Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence» (PDF). Gaia (15): 167–184.
- ^ a b Godfrey, S. J.; Altman, J. (2005). «A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon» (PDF). Jeffersoniana (16): 1–12.
- ^ STEPHEN J. GODFREY; JOHN R. NANCE; NORMAN L. RIKER (2021). «Otodus-bitten sperm whale tooth from the Neogene of the Coastal Eastern United States» (PDF). Acta Palaeontologica Polonica. 66 (3): 599–603.
- ^ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). «The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes». Journal of Mammalian Evolution. 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3. S2CID 90231.
- ^ Ehret D.J. (2010). «CHAPTER 5 – MACROEVOLUTION, AGE, AND GROWTH DETERMINATION OF THE MEGATOOTHED SHARKS (LAMNIFORMES: OTODONTIDAE)». Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes) (PDF). pp. 100–136.
- ^ S., Kenshu; Bonnan, M. F.; Becker, M. A.; Griffiths, M. L. (2021). «Ontogenetic growth pattern of the extinct megatooth shark Otodus megalodon—implications for its reproductive biology, development, and life expectancy». Historical Biology. 33 (12): 3254–3259. doi:10.1080/08912963.2020.1861608.
- ^ «Identifican en Canarias fósiles de ‘megalodón’, el tiburón más grande que ha existido» [Identifying Canary fossils of ‘megalodon’, the largest shark that ever lived] (in Spanish). Europa Press Noticias SA. 2013. Retrieved 29 August 2017.
- ^ Dulvy, N. K.; Reynolds, J. D. (1997). «Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays». Proceedings of the Royal Society B: Biological Sciences. 264 (1386): 1309–1315. Bibcode:1997RSPSB.264.1309D. doi:10.1098/rspb.1997.0181. PMC 1688595.
- ^ Godfrey, Stephen (2004). «The Ecphora» (PDF). The Newsletter of Calvert Marine Museum Fossil Club. 19 (1): 1–13. Archived from the original (PDF) on 10 December 2010.
- ^ Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (27 August 2010). «Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event». International Journal of Osteoarchaeology. 22 (3): 253–260. doi:10.1002/oa.1199.
- ^ a b Pimiento, C.; Griffin, J. N.; Clements, C. F.; Silvestro, D.; Varela, S.; Uhen, M. D.; Jaramillo, C. (2017). «The Pleistocene Marine Megafauna Extinction and its Impact on Functional Diversity». Nature Ecology and Evolution. 1 (8): 1100–1106. doi:10.1038/s41559-017-0223-6. PMID 29046566. S2CID 3639394.
- ^ a b c d «The Extinction of Megalodon». Biology of Sharks and Rays. Retrieved 31 August 2017.
- ^ Reilly, Michael (29 September 2009). «Prehistoric Shark Nursery Spawned Giants». Discovery News. Archived from the original on 10 March 2012. Retrieved 23 November 2013.
- ^ a b c Allmon, Warren D.; Steven D. Emslie; Douglas S. Jones; Gary S. Morgan (2006). «Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms». The Journal of Geology. 104 (2): 143–162. Bibcode:1996JG….104..143A. doi:10.1086/629811. S2CID 128418299.
- ^ a b Collareta, A.; Lambert, O.; Landini, W.; Bianucci, G. (2017). «Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru». Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. Bibcode:2017PPP…469…84C. doi:10.1016/j.palaeo.2017.01.001. hdl:10281/151854.
- ^ Dooly A.C.; Nicholas C. F.; Luo Z. X. (2006). «The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)». Journal of Vertebrate Paleontology. 24 (2): 453–463. doi:10.1671/2401. JSTOR 4524731. S2CID 84970052.
- ^ a b c McCormack, Jeremy; Griffiths, Michael L.; Kim, Sora L.; Shimada, Kenshu; Karnes, Molly; Maisch, Harry; Pederzani, Sarah; Bourgon, Nicolas; Jaouen, Klervia; Becker, Martin A.; Jöns, Niels (31 May 2022). «Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes». Nature Communications. 13 (1): 2980. Bibcode:2022NatCo..13.2980M. doi:10.1038/s41467-022-30528-9. ISSN 2041-1723. PMC 9156768. PMID 35641494. S2CID 249235478.
- ^ a b c d Citron, Sara; Geisler, Jonathan H.; Alberto, Collareta; Giovanni, Bianucci (2022). «Systematics, phylogeny and feeding behavior of the oldest killer whale: a reappraisal of Orcinus citoniensis (Capellini, 1883) from the Pliocene of Tuscany (Italy)». Bollettino della Società Paleontologica Italiana. 61 (2): 167–186. doi:10.4435/BSPI.2022.13.
- ^ «Huge Tooth Reveals Prehistoric Moby Dick in Melbourne». Australasian Science Magazine. Retrieved 24 April 2016.
- ^ «Move over Moby Dick: Meet Melbourne’s own mega whale». The Sydney Morning Herald. 21 April 2016.
- ^ Govender, R (2021). «Early Pliocene fossil cetaceans from Hondeklip Bay, Namaqualand, South Africa». Historical Biology. 33 (4): 574–593. doi:10.1080/08912963.2019.1650273. S2CID 202019648.
- ^ Hampe, O. (2006). «Middle/late Miocene hoplocetine sperm whale remains (Odontoceti: Physeteridae) of North Germany with an emended classification of the Hoplocetinae». Fossil Record. 9 (1): 61–86. doi:10.1002/mmng.200600002.
- ^ a b Antunes, Miguel Telles; Balbino, Ausenda Cáceres (2010). «The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic». Revista Española de Paleontología. 25 (1): 1–6.
- ^ Kast, Emma R.; Griffiths, Michael L.; Kim, Sora. L.; Rao, Zixuan C.; Shimada, Kensu; Becker, Martin A.; Maisch, Harry M.; Eagle, Robert A.; Clarke, Chelesia A.; Neumann, Allison N.; Karnes, Molly E.; Lüdecke, Tina; Leichliter, Jennifer N.; Martínez-García, Alfredo; Akhtar, Alliya A.; Wang, Xingchen T.; Haug, Gerald H.; Sigman, Daniel M. (22 June 2022). «Cenozoic megatooth sharks occupied extremely high trophic positions». Science Advances. 8 (25): eabl6529. Bibcode:2022SciA….8L6529K. doi:10.1126/sciadv.abl6529. PMC 9217088. PMID 35731884.
- ^ Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (2017). «Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics». Proceedings of the Royal Society B: Biological Sciences. 284 (1855): 20170546. doi:10.1098/rspb.2017.0546. PMC 5454272. PMID 28539520.
- ^ Sylvain, Adnet; A. C. Balbino; M. T. Antunes; J. M. Marín-Ferrer (2010). «New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean». Neues Jahrbuch für Geologie und Paläontologie. 256 (1): 7–16. doi:10.1127/0077-7749/2009/0029.
- ^ Weinstock, J. A. (2014). The Ashgate Encyclopedia of Literary and Cinematic Monsters. Farnham, United Kingdom: Routledge. pp. 107–108. ISBN 978-1-4094-2562-5. OCLC 874390267.
- ^ a b Guimont, Edward (5 October 2021). «The Megalodon: A Monster of the New Mythology». M/C Journal. 24 (5). doi:10.5204/mcj.2793. ISSN 1441-2616. S2CID 241813307.
- ^ Roesch, B. S. (1998). «A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles megalodon«. The Cryptozoology Review. 3 (2): 14–24.
- ^ «Does Megalodon Still Live?». Biology of Sharks and Rays. Retrieved 2 October 2017.
- ^ «The Meg (2018)». Box Office Mojo. Retrieved 22 December 2017.
- ^ Sid Bennett (director) (27 May 2012). Mermaids: The Body Found (Motion picture). Animal Planet.
- ^ «Shark Week ‘Megalodon: The Monster Shark Lives’ Tries To Prove Existence Of Prehistoric Shark (VIDEO)». Huff Post Green. 5 August 2013. Retrieved 11 August 2013.
- ^ Winston, B.; Vanstone, G.; Chi, W. (2017). «A Walk in the Woods». The Act of Documenting: Documentary Film in the 21st Century. New York, New York: Bloomsbury Publishing. ISBN 978-1-5013-0918-2. OCLC 961183719.
- ^ Flanagin, J. (2014). «Sorry, Fans. Discovery Has Jumped the Shark Week». New York Times. Retrieved 16 August 2014.
- ^ Shiffman, David (15 August 2014). «Shark Week Is Lying Again About Monster Megalodon Sharks». Slate Magazine. Retrieved 31 July 2022.
- ^ O’Connell, Mikey (5 August 2013). «TV Ratings: Shark Week Hits Record Highs With Fake ‘Megalodon’ Doc». The Hollywood Reporter. Retrieved 31 July 2022.
- ^ «Fossil, Fossilized Teeth of the Megalodon Shark | NCpedia». ncpedia.org. Retrieved 17 October 2019.
Further reading
- Dickson, K. A.; Graham, J. B. (November–December 2004). «Evolution and consequences of endothermy in fishes». Physiological and Biochemical Zoology. 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772. S2CID 40104003.
- Kent, Bretton W. (1994). Fossil Sharks of the Chesapeake Bay Region. Columbia, Md.: Egan Rees & Boyer. ISBN 978-1-881620-01-3. OCLC 918266672.
External links
- The rise of super predatory sharks
- Carcharocles: Extinct Megatoothed shark
- Paleontological videos
- Paleontologist Mark Renz shows one of the largest megalodon teeth discovered on YouTube
- Shark Week Special on megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about megalodon on YouTube (featuring expert Dana Ehret)
- Lamniform sharks: 110 million years of ocean supremacy on YouTube (featuring expert Mikael Siverson)
- The Rise and Fall of the Neogene Giant Sharks on YouTube (featuring expert Bretton Kent)
- Moore, Kallie (19 December 2018). «Why Megalodon (Definitely) Went Extinct». PBS Eons. Archived from the original on 14 November 2021 – via YouTube.
Вымершие виды гигантских акул от 23 до 3,6 миллиона лет назад
| Мегалодон. Временной диапазон: Бурдигал — Занклин, ок. 23–3.6 Ma PreꞒ Ꞓ O S D C P T J K Pg N | |
|---|---|
|
|
| Модель челюстей мегалодона в Американском музее естественной истории | |
| Научная классификация | |
| Домен: | Eukaryota |
| Королевство: | Animalia |
| Тип: | Chordata |
| Класс: | Chondrichthyes |
| Отряд: | Lamniformes |
| Семейство: | †Otodontidae |
| Род: | †Otodus |
| Вид: | †О. megalodon |
| Биномиальное имя | |
| † Otodus megalodon . (Agassiz, 1843) | |
| Синонимы | |
Список синонимов
|
Megalodon (Otodus megalodon), что означает «большой зуб», является вымершим видом из акул, который жил примерно от 23 до 3,6 миллионов лет назад (млн лет назад), в период раннего миоцена до плиоцена. Ранее считалось, что это член семейства Lamnidae и близкий родственник большой белой акулы (Carcharodon carcharias). Однако в настоящее время они классифицируются как вымершее семейство Otodontidae, которое отделились от большой белой акулы в раннемеловом. Его родовое положение до сих пор обсуждается, авторы помещают его либо в Carcharocles, ни в Megaselachus, ни в Otodus, ни в Procarcharodon. Это связано с тем, что были обнаружены переходные окаменелости, показывающие, что мегалодон является последним хроновидом линии гигантских акул, первоначально принадлежавших к роду Otodus, который возник в палеоцене.
. Считающийся одним из самых крупных и могущественных хищников, когда-либо существовавших, мегалодон известен по фрагментарным останкам, и его внешний вид и максимальный размер неизвестны. Ученые расходятся во мнениях относительно того, напоминала бы она более крупную версию большой белой акулы, китовой акулы (Rhincodon typus), гигантской акулы (Cetorhinus maximus) или песчаная тигровая акула (Carcharias taurus). Большинство оценок размера мегалодона основаны на зубах; с максимальной оценкой длины до 14,2–16 метров (47–52 футов) и средней оценкой длины 10,5 метра (34 фута). По оценкам, их большие челюсти могут оказывать силу укуса от 108 500 до 182 200 ньютонов (от 24 400 до 41 000 фунтов силы). Их зубы были толстыми и крепкими, чтобы хватать добычу и ломать кости.
Мегалодон, вероятно, оказал большое влияние на структуру морских сообществ. Летопись окаменелостей указывает на то, что он имел космополитическое распространение. Вероятно, он был нацелен на крупную добычу, такую как киты, тюлени и морские черепахи. Молодь населяла теплые прибрежные воды и питалась рыбой и мелкими китами. В отличие от большого белого, который атакует добычу с мягкой нижней стороны, мегалодон, вероятно, использовал свои сильные челюсти, чтобы пробить грудную полость и проколоть сердце и легкие своей добычи.
Животное столкнулось с конкуренцией со стороны китообразных, питающихся китами, таких как Ливятан и других макрорапториальных кашалотов и, возможно, более мелких предков-косаток. Поскольку акула предпочитала более теплые воды, считается, что охлаждение океана, связанное с наступлением ледниковых периодов, в сочетании с понижением уровня моря и, как следствие, потерей подходящих зон для рассадников, также могло способствовать его сокращению.. Уменьшение разнообразия усатых китов и смещение их распределения в сторону полярных регионов, возможно, уменьшили основной источник пищи мегалодона. Вымирание акул, по-видимому, затронуло и других животных; например, размер усатых китов значительно увеличился после того, как акула исчезла.
Содержание
- 1 Таксономия
- 1.1 Именование
- 1.2 Эволюция
- 2 Биология
- 2.1 Внешний вид
- 2.2 Размер
- 2.2.1 Средние оценки
- 2.2.2 Максимальные оценки
- 2.3 Зубы и сила укуса
- 2.4 Внутренняя анатомия
- 3 Палеобиология
- 3.1 Ареал и среда обитания
- 3.1.1 Местонахождение окаменелостей
- 3.2 Отношения между жертвами
- 3.3 Конкуренция
- 3.4 Стратегии кормления
- 3.5 Ясли
- 3.1 Ареал и среда обитания
- 4 Вымирание
- 4.1 Изменение климата
- 4.2 Изменение экосистемы
- 5 В массовой культуре
- 6 См. Также
- 7 Примечания
- 8 Ссылки
- 9 Дополнительная литература
- 10 Внешние ссылки
Таксономия
Именование
Изображение головы акулы Николасом Стено в его работе «Голова акулы иссечена»
Согласно отчетам эпохи Возрождения, гигантские треугольные окаменелые зубы, часто обнаруживаемые в скалистых образованиях, когда-то считались окаменевшими языками, или glossopetrae, драконов и змеи. Это толкование было исправлено в 1667 году датским натуралистом Николасом Стено, который узнал в них зубы акулы и, как известно, создал изображение акулы Голова несёт такие зубы. Он описал свои находки в книге «Голова рассеченной акулы», которая также содержала иллюстрацию зуба мегалодона.
Швейцарский натуралист Луи Агассис дал этой акуле начальный научное имя, Carcharodon megalodon, в его работе 1843 года Recherches sur les poissons fossiles, основанной на остатках зубов. Английский палеонтолог Эдвард Чарльзуорт в своей статье 1837 года использовал имя Carcharias megalodon, цитируя Агассиса как автора, что указывает на то, что Агассис описал этот вид до 1843 года. Английский палеонтолог Чарльз Дэвис Шерборн в 1928 году перечислил серию статей Агассиса за 1835 год как первое научное описание акулы. конкретное имя мегалодон переводится как «большой зуб», от древнегреческого : μέγας, латинизировано : (мегас), лит. ‘большой, могучий’ и ὀδούς (odoús), «зуб». Зубы мегалодона морфологически сходны с зубами большой белой акулы (Carcharodon carcharias), и на основании этого наблюдения Агассис отнес мегалодона к роду Carcharodon. Хотя «мегалодон» — неофициальное название акулы, ее также часто неофициально называют «гигантская белая акула», «мегалодонная акула», «большая зубная акула» или «Мэг».
Там было одним из очевидных описаний акулы в 1881 г., классифицирующим ее как Selache manzonii.
Evolution
| Взаимоотношения между мегалодоном и другими акулами, в том числе большой белой акулой (Carcharodon carcharias) |
Хотя самые ранние останки мегалодона были зарегистрированы в позднем олигоцене, примерно 28 миллионов лет назад назад (млн лет назад), существуют разногласия относительно того, когда оно появилось, датируется всего 16 млн лет назад. Считалось, что мегалодон вымер примерно в конце плиоцена, около 2,6 млн лет назад; утверждения о зубах плейстоцена мегалодона, возраст которых меньше 2,6 миллиона лет, считаются ненадежными. Оценка 2019 года переносит дату вымирания на более ранний период плиоцена, 3,6 млн лет назад.
Мегалодон теперь считается членом семейства Otodontidae, рода Otodus, в отличие от его предыдущей классификации в Lamnidae, род Carcharodon. Отнесение мегалодона к Carcharodon было обусловлено сходством зубов с большой белой акулой, но большинство авторов в настоящее время считают, что это связано с конвергентной эволюцией. В этой модели большая белая акула более тесно связана с вымершим широкозубым мако (Isurus hastalis ), чем с мегалодоном, о чем свидетельствуют более похожие зубы у этих двух акул; Зубы мегалодона имеют гораздо более тонкие зазубрины, чем зубы большой белой акулы. Большая белая акула более тесно связана с акулой мако (Isurus spp.), Имеющей общего предка около 4 млн лет назад. Сторонники первой модели, в которой мегалодон и большая белая акула более тесно связаны между собой, утверждают, что различия между их зубами незначительны и неясны.
Зуб мегалодона с двумя зубами большой белой акулы
В настоящее время род Carcharocles насчитывает четыре вида: C. auriculatus, C. angustidens, C. chubutensis и C. megalodon. Эволюция этой линии характеризуется увеличением зазубрин, расширением коронки, развитием более треугольной формы и исчезновением боковых бугров. Эволюция морфологии зубов отражает сдвиг в тактике хищничества от рвущего и захватывающего укусов к режущему, что, вероятно, отражает сдвиг в выборе добычи от рыб к китообразным. Боковые бугра окончательно утрачены в процессе постепенного перехода от C. chubutensis к C. megalodon, который длился примерно 12 миллионов лет. Этот род был предложен Д. С. Джорданом и Х. Ганнибалом в 1923 г. для содержания C. auriculatus. В 1980-х годах мегалодон был отнесен к Кархароклу. До этого, в 1960 году, французский ихтиолог Эдгар Казье установил род Procarcharodon, который включал в себя этих четырех акул и считался отдельным от большой белой акулы. Сейчас он считается младшим синонимом Кархарокла. Род Palaeocarcharodon был возведен рядом с Procarcharodon, чтобы представить начало рода, а в модели, в которой мегалодон и большая белая акула тесно связаны, их последний общий предок. Авторы, отвергающие эту модель, считают его тупиком эволюции и не имеют отношения к акулам Carcharocles.
большая белая акула (Carcharodon carcharias) и мегалодон ранее были считаются близкими родственниками.
Другая модель эволюции этого рода, также предложенная Касье в 1960 году, состоит в том, что прямым предком кархароклов является акула Otodus obliquus, которая жила с Палеоцен — миоцен эпохи, 60–13 млн лет назад. Род Otodus произошел от Cretolamna, акулы мелового периода. В этой модели O. obliquus превратился в O. aksuaticus, который превратился в C. auriculatus, затем в C. angustidens, затем в C. chubutensis, а затем, наконец, в C. megalodon.
Другая модель эволюции кархарокла, предложенная в 2001 году палеонтологом Майклом Бентоном, состоит в том, что три других вида на самом деле представляют собой единый вид акул, который постепенно менялся с течением времени между палеоценом и плиоцен, что делает его хроновидом. Некоторые авторы предполагают, что C. auriculatus, C. angustidens и C. chubutensis следует классифицировать как один вид в роду Otodus, оставив C. megalodon единственным представителем Carcharocles.
Род Carcharocles может быть недействительным., и акула может действительно принадлежать к роду Otodus, что делает ее Otodus megalodon. Исследование палеогеновых акул 1974 года, проведенное Анри Каппеттой, выявило подрод Megaselachus, классифицируя акулу как мегалодона Otodus (Megaselachus) вместе с O. (M.) chubutensis. Обзор Chondrichthyes в 2006 году повысил Megaselachus до рода и классифицировал акул как Megaselachus megalodon и M. chubutensis. Обнаружение окаменелостей, отнесенных к роду Megalolamna в 2016 году, привело к переоценке Otodus, в результате которой был сделан вывод, что он парафилетический, то есть состоит из последних общий предок, но он не включает всех его потомков. Включение акул Кархарокла в Отодус сделало бы его монофилетическим, а сестринской кладой является мегалоламна.
Биология
Внешний вид
Восстановление предполагая внешнее сходство с большой белой акулой
Одна из интерпретаций того, как появился мегалодон, заключалась в том, что это была крепкая акула и, возможно, имела сходное телосложение с большой белой акулой. Челюсти могли быть более тупыми и шире, чем у большого белого, и плавники также были бы похожи по форме, хотя и толще из-за своего размера. Возможно, у него был косоглазый вид, поскольку у него были маленькие глубоко посаженные глаза.
Другая интерпретация заключается в том, что мегалодон имел сходство с китовой акулой (Rhincodon typus) или гигантская акула (Cetorhinus maximus). Хвостовой плавник имел бы форму полумесяца, анальный плавник и второй спинной плавник были бы небольшими, и по обе стороны от хвостового плавника был бы хвостовой киль (на хвостовой стебель ). Эта конструкция характерна для других крупных водных животных, таких как киты, тунцы и другие акулы, для уменьшения сопротивления во время плавания. Форма головы может варьироваться у разных видов, так как большинство адаптаций, снижающих сопротивление, приходится на хвост животного.
Скульптура в Музее эволюции в Пуэбле, Мексика
Так как Кархарокл произошел от Отодуса, и у двоих были зубы, которые очень похожи на зубы песчаной тигровой акулы (Carcharias taurus), мегалодон мог иметь телосложение, более похожее на песчаную тигровую акулу, чем на других акул. Это маловероятно, поскольку песчаная тигровая акула — пловец в форме черепах, которому для движения по воде требуется более быстрое движение хвоста, чем большая белая акула, пловец в форме грозы.
Размер
Средние оценки
Сравнение размеров большой белой и китовой акулы с оценками мегалодона Пропорции мегалодона при длине 3 м (9,8 фута), 8 м (26 футов) и 16 м (52 фута), экстраполированное от ныне живущих родственников, с дайвером длиной 1,65 м (5 футов 5 дюймов)
Из-за фрагментарных останков было получено много противоречивых оценок размеров мегалодона, поскольку они могут быть получены только по ископаемым зубам и позвонкам. Также из-за этого большая белая акула является основой ее реконструкции и оценки размеров, так как считается лучшим аналогом мегалодона. Используя оценки длины, экстраполированные на основе 544 зубов, найденных за все геологическое время и географию, включая взрослых и молодых, исследование 2015 года оценило среднюю длину в 10,5 метров (34 фута). Для сравнения: максимальный зарегистрированный размер большой белой акулы составляет 6,1 метра (20 футов), а китовая акула (самая большая из ныне живущих рыб) может достигать 18,8 метра (62 фута). Возможно, что разные популяции мегалодонов по всему миру имели разные размеры тела и поведение из-за разного экологического давления. В любом случае считается, что это была самая крупная хищная акула из когда-либо существовавших. Исследование 2020 года, изучившее размеры современных больших белых акул, мако и ламна, показало, что мегалодон длиной 16 м (52 фута) имел бы длину головы 4,65 м (15,3 фута), 1,41 м. (4 фута 8 дюймов) высокие жаберные щели, высокий спинной плавник 1,62 м (5 футов 4 дюйма), длина грудных плавников 3,08 м (10 футов 1 дюйм) и хвостовой плавник высотой 3,85 м (12 футов 8 дюймов).
Зрелый самец мегалодона мог иметь массу от 12,6 до 33,9 метрических тонн (от 13,9 до 37,4 коротких тонн), а зрелые самки — от 27,4 до 59,4 метрических тонн (от 30,2 до 65,5 коротких тонн), если предположить, что самцы могла иметь длину от 10,5 до 14,3 метра (от 34 до 47 футов), а женщины — от 13,3 до 17 метров (от 44 до 56 футов). Исследование 2015 года, связывающее размер акулы и типичную скорость плавания, показало, что мегалодон обычно плавает со скоростью 18 километров в час (11 миль в час) — при условии, что его масса тела обычно составляла 48 метрических тонн (53 коротких тонны), что согласуется с другими водными существами. таких размеров, как финват (Balaenoptera Physalus), который обычно курсирует со скоростью от 14,5 до 21,5 км / ч (от 9,0 до 13,4 миль в час).
Его большие размеры могли быть из-за климатических факторов и обилия крупных объектов добычи, а также, возможно, на это повлияла эволюция региональной эндотермии (мезотермия), которая увеличила бы скорость метаболизма и скорость плавания. Считалось, что акулы-отодонтиды были эктотермными, так что на этом основании мегалодон мог быть эктотермным. Однако крупнейшие современные эктотермические акулы, такие как китовая акула, являются фильтраторами, а ламниды, как теперь известно, являются региональными эндотермами, что подразумевает некоторые метаболические корреляции с хищным образом жизни. Эти соображения, а также данные об изотопах кислорода в зубах и необходимость более высоких скоростей всплеска плавания у макропохищников эндотермической добычи, чем позволяет эктотермия, подразумевают, что отодонтиды, включая мегалодона, вероятно, были региональными эндотермами.
Максимальные оценки
Гордон Хаббелл из Гейнсвилля, Флорида обладает верхним передним зубом мегалодона, максимальная высота которого составляет 18,4 см (7,25 дюйма), это один из крупнейших известных образцов зубов акулы. Кроме того, реконструкция челюсти мегалодона размером 2,7 на 3,4 метра (9 на 11 футов), разработанная охотником за окаменелостями Вито Бертуччи, содержит зуб, максимальная высота которого, как сообщается, превышает 18 сантиметров (7 дюймов).
Первый Попытка восстановить челюсть мегалодона была предпринята Башфордом Дином в 1909 году и выставлена в Американском музее естественной истории. Исходя из размеров этой реконструкции челюсти, было выдвинуто предположение, что мегалодон мог достигать 30 метров (98 футов) в длину. Дин переоценил размер хряща на обеих челюстях, из-за чего он оказался слишком высоким.
Реконструкция Башфорда Дина в 1909 году Зуб по сравнению с рукой
В 1973 году Джон Э. Рэндалл, ихтиолог, использовал высоту эмали (вертикальное расстояние лезвия от основания эмалевой части зуба до его кончика) для измерения длины акулы. давая максимальную длину около 13 метров (43 фута). Однако высота зубной эмали не обязательно увеличивается пропорционально общей длине животного.
В 1994 году морские биологи Патрик Дж. Шембри и Стивен Папсонпредположили, что О. мегалодон, возможно, приблизился к максимуму примерно от 24 до 25. метров (от 79 до 82 футов) общей длиной.
В 1996 году исследователи акул Майкл Д. Готфрид, Леонард Компаньо и С. Кертис Боуман предложили линейную зависимость между общей длиной акулы. и высота самого большого переднего зуба в верхней части. Предлагаемое соотношение: общая длина в метрах = — (0,096) × [максимальная высота UA (мм )] — (0,22). Они утверждали, что общая длина О. мегалодона могла достичь не более 20,3 метра (67 футов).
В 2002 году исследователь акул Клиффорд Джеремайя предположил, что общая длина ширине корня переднего переднего зуба.. Он утверждал, что на каждый 1 сантиметр (0,39 дюйма) ширины корня приходится примерно 1,4 метра (4,6 фута) длины акулы. Инструмент для оценки периметра челюсти является инструментом оценки периметра самых больших зубов. Самый большой зуб, которому владел Иеремия, имел длину корня около 12 сантиметров (4,7 дюйма), что давало общую длину 16,5 метра (54 фута).
В 2002 году палеонтолог Кеншу Шимада из Университета ДеПол предложили линейную зависимость между высотой коронки зуба и общей длины после проведения анатомического анализа нескольких образцов, что можно использовать зуб любого размера. Шимада заявил, что ранее предложенные методы были основаны на менее надежной оценке стоматологической гомологии между мегалодоном и большой белой акулой, и что скорость роста между короной и корнем не изометрической. модели. Используя эту модель, верхний передний зуб Готфрида и его коллега имела общую длину 15 метров (49 футов). Среди нескольких образцов, найденных в форме Гатун в Панаме, один верхний боковой зуб был использован другими исследователями для использования общей оценки длины 17,9 метра (59 футов) с помощью этого метода.
В 2019 году Шимада пересмотрел мегалодона и не рекомендуется использовать не передние зубы для оценки, отмечая, что размер положение положение не передних зубов определить трудно. Симада заявленная максимальная общая длина оценок, основанная на верхних передних зубах, которые доступны в музеях, составляет 14,2 и 15,3 метра (47 и 50 футов), в зависимости от используемого метода оценки.
Зубы и сила прикуса
Реконструкция, показывающая положение замещающих зубов
Наиболее распространенными окаменелостями мегалодона являются его зубы. Диагностические характеристики включают треугольную форму, прочную структуру, большой размер, мелкие зазубрины, отсутствие боковых зубцов и видимую V-образную шейку (где корень соответствует корона ). Зуб встретился с челюстью под крутым углом, как у большой белой акулы. Зуб был закреплен с помощью волоконной соединительной ткани, и шероховатость основания, возможно, добавляла механической прочности. Язычная сторона зуба, обращенная к языку часть, была выпуклой; а губная сторона, другая сторона зуба, была слегка выпуклой или плоской. Передние зубы были почти перпендикулярны челюсти и симметричны, тогда как задние зубы были наклонными и асимметричными.
Зубы мегалодона могут иметь наклонную высоту (длина диагонали) более 180 миллиметров (7,1 дюйма) и являются высокими из них. любой известный вид акул, подразумевая, что это была самая большая из всех хищных акул. В 1989 г. почти полный набор зубов мегалодона был обнаружен в Сайтаме, Япония. Другой почти полный зубной мегалодона был извлечен из Йорктаунских формаций в США и использовался рядил для реконструкции челюсти мегалодона в Национальном музее естественной истории (USNM). На основе этих открытий в 1996 году для мегалодона была создана искусственная зубная формула.
Зубная формула мегалодона: 2.1.7.43.0.8.4. Как видно из формулы, у мегалодона в челюстях было четыре вида зубов: передние, промежуточные, боковые и задние. Промежуточный зуб мегалодона технически выглядит как верхний передний и обозначается как «A3», потому что он довольно симметричен и не направлен мезиально (сторона зуба к средней линии челюстей, где левая и правая челюсти встретить). У мегалодона были очень крепкие зубные ряды, и в его челюстях было более 250 зубов, охватывающих 5 рядов. Возможно, челюсти крупных мегалодонов составляли примерно 2 метра в поперечнике. Зубы также были зазубренными, что повысило эффективность прорезания плоти или кости. Акула могла открывать пасть под углом 75 °, реконструкция в USNM составляет примерно 100 °.
Реконструированные челюсти выставлены в Национальном аквариуме в Балтиморе
В 2008 г. Группа ученых под руководством С. Рое провела эксперимент по определению силы укуса большой белой акулы, используя образец длиной 2,5 метра (8,2 фута), а изометрически масштабировала результаты до максимума. размер и консервативный минимум и максимум массы тела мегалодона. Они поместили силу укуса последней от 108 514 до 182 201 ньютона (от 24 395 до 40 960 фунтов силы) в задний прикус, по сравнению с силой укуса 18 216 ньютонов (4095 фунтов силы) для самой большой подтвержденной большой белой акулы и 7400 ньютонов ( 1700 фунтов силы). для плакодермы рыбы Dunkleosteus. Кроме того, вро и его коллеги отметили, что акулы трясутся боком во время кормления, усиливая создаваемую силу, что, вероятно, достигло того, что общая сила, испытываемая добыча, была выше расчетной.
Внутренняя анатомия
Реконструировано Скелет мегалодона, представленный в Морском музее Калверта
Мегалодон представлен в летописи окаменелостей зубами, центрами позвонков и копролитами. Как и у всех акул, скелет мегалодона был сформирован из хряща, а не кости ; следовательно, большинство ископаемых плохо образцов сохранились. Чтобы поддерживать большой зубной ряд, челюсти мегалодона были бы более массивными, крепкими и развитыми, чем у большого белого человека, который обладает сравнительно изящными зубными рядами. Его хондрокраниум, хрящевой череп, имел бы более массивный и массивный вид, чем у большого белого. Его плавники были пропорциональны его большему размеру.
Было найдено несколько ископаемых позвонков. Наиболее примечательным примером является частично сохранившийся позвоночный столб одного образца, раскопанный в Антверпенской котловине, Бельгия, в 1926 году. Он включает 150 центральных, причем центральные части находятся в диапазоне от 55 от миллиметров (2,2 дюйма) до 155 миллиметров (6 дюймов) в диаметре. Позвонки акулы, возможно, стали намного больше, и изучение экземпляров показало, что у него было больше позвонков, чем у экземпляров любой известной акулы, возможно, более 200 центров; только великий приблизился к нему. Другой частично сохранившийся позвоночный столб мегалодона был раскопан в форме Грам в Дании в 1983 году, он включает 20 центральных частей позвоночного с диаметром от 100 миллиметров (4 дюйма) до 230 миллиметров (9 дюймов).
Копролит приписывается мегалодону
Копролитовые остатки мегалодона имеют спиралевидную форму, что указывает на то, что у акулы мог быть спиральный клапан, часть в форме штопора нижний отделов кишечника, похожи на современные пластинчатых акул. Остатки миоценового копролита были обнаружены в округе Бофорт, Южная Каролина, причем один из них был размером 14 см (5,5 дюйма).
Готфрид и его коллеги реконструировали весь скелет мегалодона, который позже был выставлен на обозревателе Морском музее Калверта в США и Южноафриканском музее Изико. Эта реконструкция имеет длину 11,3 метра (37 футов) и представляет зрелого самца на основе онтогенетических изменений, которые пережила большая белая акула в течение жизни.
Палеобиология
Ареал и среда обитания
Мегалодон имел космополитическое распространение ; его окаменелости были раскопаны во многих частях мира, включая Европу, Африку, Америку и Австралию. Чаще всего это происходило в субтропических и умеренных широтах. Он был обнаружен на широтах до 55 ° с.ш. ; его предполагаемый допустимый диапазон температур составляющий 1-24 ° C (34-75 ° F). Вероятно, он обладает способностью переносить такие низкие температуры из-за мезотермии, физиологические способности организма акул активации метаболическое тепло, поддерживая более высокую температуру тела, чем окружающая вода.
Мегалодон населял широкий спектр морской среды (например, мелководные прибрежные воды, районы прибрежного апвеллинга, заболоченные прибрежные лагуны, песчаные литорали и прибрежные глубоководные среды) и показал преходящий образ жизни. Взрослые мегалодоны не были многочисленны на мелководье и в основном населяли прибрежные районы. Мегалодон мог перемещаться между прибрежными и океаническими водами, особенно на разных стадиях своего жизненного цикла.
Ископаемые останки тенденцию к тому, что образцы в южном полушарии в среднем больше, чем в северном, со средней длиной 11,6 и 9,6 метра (38 и 31 фут) соответственно; и также больше в Тихом океане, чем в Атлантике, со средней длиной 10,9 и 9,5 метра (36 и 31 фут) соответственно. Они не предполагают какой-либо тенденции к изменению размера с абсолютной широтой или к изменению размера с течением времени (хотя в целом считается, что происхождение Carcharocles демонстрирует тенденцию к увеличению размера с течением времени). Общая модального оценивается в 10,5 метров (34 фута), причем распределение длин смещено в сторону более крупных особей, что свидетельствует об экологическом или конкурентном преимуществе для большего размера тела.
Расположение окаменелостей
Мегалодон имел глобальное распространение, и окаменелости акулы были найдены во многих местах по всему миру, омывающих все океаны неогена.
| эпохи | формации | Государство | Континент |
|---|---|---|---|
| Плиоцен | Африка | ||
| Африка | |||
| Африка | |||
| Европа | |||
| Европа | |||
| Европа | |||
| Европа | |||
| Формация Красного Скала | Европа | ||
| Формация Сан-Матео | Северная Америка | ||
| Формация Таусли | Северная Америка | ||
| Формация Боун-Вэлли | Северная Америка | ||
| Формация Тамиами | Северная Америка | ||
| Йорктаунская формация | Северная Америка | ||
| Формация Холмов | Северная Америка | ||
| Северная Америка | |||
| Формация Сан-Диего | Северная Америка | ||
| Формация Тирабузон | Севе р Америка | ||
| Формация Онзоле | Южная Америка | ||
| Южная Америка | |||
| Океания | |||
| Океания | |||
| Океания | |||
| Океания | |||
| Океания | |||
| Океания | |||
| Миоцен | |||
| Африка | |||
| Африка | |||
| Африка | |||
| Африка | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Азия | |||
| Европа | |||
| Формация Мелкер-Санд | Европа | ||
| Европа | |||
| Формация Вайссенегг | Европа | ||
| Европа | |||
| Европа | |||
| Европа | |||
| Грамм-образование | Европа | ||
| Бассейн Аквитании | Европа | ||
| Европа | |||
| Европа | |||
| Формация голубой глины | Европа | ||
| Известняк Глобигерина | Европа | ||
| Европа | |||
| Формация Бреда | Европа | ||
| Европа | |||
| Известняк Leitha | Европа | ||
| Европа | |||
| Европа | |||
| Европа | |||
| Европа | |||
| Европа | |||
| формация Монтерей | Северная Америка | ||
| Формация Пуэнте | Северная Америка | ||
| Формация Пурисима | Северная Америка | ||
| Формация Сан-Матео | Северная Америка | ||
| Формация Санта-Маргарита | Северная Америка | ||
| Формация Темблор | Северная Америка | ||
| Формация Топанга | Северная Америка | ||
| Формация Bone Valley | Северная Америка | ||
| Формация Калверт | Северная Америка | ||
| Северная Америка | |||
| Северная Америка | |||
| Северная Америка | |||
| Северная Америка | |||
| Северная Америка | |||
| Известняк Аймамон | Северная Америка | ||
| Формация Альмехас | Северная Америка | ||
| Формация Каррильо-Пуэрто | Северная Америка | ||
| Формация Чагрес | Северная Америка | ||
| Северная Америка | |||
| Формация Гатун | Северная Америка | ||
| Южная Америка | |||
| Формация Баия Инглеса | Южная Америка | ||
| Формация Кастильетес | Южная Америка | ||
| Южная Америка | |||
| Формация Писко | Южная Америка | ||
| Южная Америка | |||
| Южная Америка | |||
| Южная Америка | |||
| Южная Америка | |||
| Формация Урумако | Южная Америка | ||
| Океания | |||
| Океания | |||
| Океания | |||
| Формация Маннум | Океания | ||
| Океания | |||
| Ок. eania | |||
| Океания | |||
| Океания |

![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() Места обнаружения окаменелостей мегалодонов, желтые из плиоцена и синие из миоцена
Места обнаружения окаменелостей мегалодонов, желтые из плиоцена и синие из миоцена
Отношения с добычей
Позвонок кит, укушенный пополам мегалодоном, с видимыми порезами на зубах
Хотя акулы, как правило, приспособлены для кормления, большие размеры мегалодона, способность быстро плавать и мощные челюсти в сочетании с впечатляющим устройством для кормления сделали его вершиной хищник, способный поедать широкий спектр животных. Вероятно, это был один из самых сильных хищников на свете. Исследование, посвященное изотопам кальция вымерших и современных эластожаберных акул и скатов, показало, что мегалодон питался на более высоком трофическом уровне, чем современная большая белая акула. То есть он был выше в пищевой цепи.
. Ископаемые свидетельства указывают на то, что мегалодон охотился на многих видов китообразных, таких как дельфины, мелкие киты, цетотеры, сквалодонтиды (зубастые дельфины), кашалоты, гренландские киты и рорквалы. В дополнение к этому они также предназначались для тюленей, сирен и морских черепах. Акула была оппортунистом и рыбоядным, и она также должна была преследовать более мелкую рыбу и других акул. На многих китах были обнаружены глубокие порезы, скорее всего, сделанные их зубами. При различных раскопках были обнаружены зубы мегалодона, лежащие рядом с пережеванными останками китов, а иногда и в прямой связи с ними.
Экология питания мегалодона, по-видимому, менялась в зависимости от возраста и между участками, как и у современной. большая белая птицы. Вполне вероятно, что популяция взрослых мегалодонов у побережья Перу в первую очередь предназначалась для цетотерских китов длиной от 2,5 до 7 метров (от 8,2 до 23 футов) и другой доход меньше, чем она сама, а не для крупных китов того же класса, что и они сами. Между тем, у молодых особей, вероятно, было больше рыбы.
Конкуренция
Мегалодон мог столкнуться с конкуренцией со стороны макропториальных кашалотов, таких как Ливятан (вверху).
Мегалодон столкнулся с конкурентной средой. Его положение на вершине пищевой цепи, вероятно, оказало значительное влияние на структурирование морских сообществ. Ископаемые остатки указывают на корреляцию между мегалодоном и появлением и диверсификацией китообразных и других морских млекопитающих. Мегалодоны предпочитают места обитания, в которых было много мелких китообразных, а взрослые. Мегалодоны предпочитают места обитания, в которых были изобилии крупные китообразные. Такие предпочтения могли развиться вскоре после того, как они появились в олигоцене.
Megalodon были современниками китоядных зубатых китов (особенно макрорапториальных кашалотов и сквалодонтид), которые, вероятно, также были одними из главных хищников эпохи и обеспечлицию. Некоторые достигли гигантских размеров, например Ливятан, который вырос с 13,5 до 17,5 метров (с 44 до 57 футов). Окаменелые зубы неопределенного вида таких фитероидов из рудника Ли-Крик, Северная Каролина, указать на то, что максимальная длина тела у них составляла 8–10 м, максимальная продолжительность жизни составляла около 25 лет. Это сильно отличается от современных косаток аналогичного размера, которые позволяют предположить, что это отличие от последних, которые являются высшими хищниками, эти физетероиды подверглись хищничеству со стороны более крупных видов, таких как мегалодон или ливятан. К позднему миоцену, примерно к 11 млн лет назад, численность и разнообразие макрорапториалов резко упали. Другие виды, возможно, заполнили эту нишу в плиоцене, например, ископаемый косатка Orcinus citoniensis, который мог быть стайным хищником и преследовать жертву крупнее, чем он сам, но этот вывод оспаривается, и, вероятно, это был хищник-универсал, а не специалист по морским млекопитающим.
Мегалодон мог подвергать современных белых акул конкурентному исключению, поскольку ископаемые виды показывают, что другие акул избегали регионов, где он населен, в основном придерживаясь более холодных вод того времени. В областях, где их ареалы, казалось, перекрывались, например, в плиоцене Нижняя Калифорния, возможно, что мегалодон и большая белая акула населяли этот район в разное время года, преследуя разную мигрирующую добычу. Мегалодон, вероятно, имеет склонность к каннибализму, как и современные акулы.
Стратегии кормления
Художественное впечатление мегалодона, преследующего двух Eobalaenoptera китов
акул часто используют сложные стратегии охоты для охоты на больших хищников. Стратегия охоты на большую белую акулу может быть похожа на то, как мегалодон охотился на свою крупную добычу. Следы укусов мегалодона на окаменеллах кита позволяют предположить, что он использовал разные стратегии охоты против крупной производительности, чем большая белая акула.
Один особый образец — останки неописанного миоценового усатого кита длиной 9 метров (30 футов) — предоставлены первая возможность количественно проанализировать его атакующее поведение. В отличие от больших белых, которые нацелены на нижнюю часть живота своей добычи, мегалодон, вероятно, нацелился на сердце и легкие, с их толстыми зубами, приспособленными для прокалывания твердых костей, о чем свидетельствуют следы укусов, нанесенные на грудную клетку и другие жесткие костные области на останках китов. Кроме того, модели атаки могут отличаться для добычи разного размера. Ископаемые останки некоторых мелких китообразных, например цетотериев, предполагают, что они были тараны снизу с большой силой, прежде чем их убили и съели, на основании компрессионных переломов.
В плиоцене появились более крупные китообразные. Megalodon, по-видимому, усовершенствовал свои стратегии охоты, чтобы справиться с этими большими китами. Многочисленные окаменелые кости плавников и позвонки хвоста крупных китов из плиоцена были обнаружены со следами укусов мегалодона, что позволяет предположить, что мегалодон обездвижил крупного кита перед тем, как убить его и поесть. 455>
Питомники
Коллекция зубов молодых мегалодонов и C. chubutensis из вероятного питомника в формации Gatún в Панаме
Мегалодон, как и современные акулы, использовали места для выращивания своих детенышей, особенно в теплой прибрежной среде с большим количеством еды и защиты от хищников. Питомники были обнаружены в формации Гатун в Панаме, в формации Калверт в Мэриленде, Банко-де-Консепсьон на Канарских островах и в формации Кости-Вэлли во Флориде.. Учитывая, что все дошедшие до нас пластинчатые акулы рождают живых детенышей, считается, что это относится и к мегалодонам. Младенцы-мегалодоны были около 3,5 метров (11 футов) в своем наименьшем размере, а детеныши были уязвимы для хищников других видов акул, таких как большая акула-молот (Sphyrna mokarran) и зубастая акула (Hemipristis serra). Их диетические предпочтения демонстрируют онтогенетический сдвиг: молодые мегалодоны обычно охотятся на рыбу, морских черепах, дюгоней и мелких китообразных; зрелый мегалодон переместился в прибрежные районы и поедал крупных китообразных.
Исключительный случай в летописи окаменелостей предполагает, что молодой мегалодон мог иногда нападать на гораздо более крупных балаеноптерид китов. Три следа зубов, по-видимому, от 4-7 метровой (13-23 футов) акулы плиоцена были найдены на ребре синего или горбатого кита, являющегося предком, что свидетельствовало о последующем заживлении, которое, как предполагается, было нанесено молодой мегалодон.
Исчезновение
Изменение климата
Земля испытала ряд изменений в течение периода существования мегалодона, которые повлияли на морскую жизнь. Тенденция похолодания, начавшаяся в олигоцене 35 млн лет назад, в конечном итоге привела к оледенению на полюсах. Геологические события изменили течения и осадки; Среди них были закрытие Центральноамериканского морского пути и изменения в океана Тетис, способствовавшие охлаждению океанов. Из-за остановки Гольфстрима вода, богатая питательными веществами, не попала в основные морские экосистемы, что могло негативно повлиять на ее источники питания. Наибольшее колебание уровня моря в кайнозойской эре произошло в плио-плейстоцене, примерно между 5 миллионами и 12 тысячами лет назад, из-за расширения ледников на полюсах, которые отрицательно повлияло на прибрежную среду и, возможно, способствовало его исчезновению вместе с таковыми нескольких других морских видов мегафауны. Эти океанографические изменения, в частности, падение уровня моря, возможно, ограничили многие подходящие мелководные теплые места рассадников мегалодона, препятствуя их размножению. Районы питомников имеют решающее значение для выживания многих видов акул, отчасти потому, что они защищают молодь от хищников.
Поскольку его ареал явно не распространялся на более холодные воды, мегалодон, возможно, не смог удержать значительное количество метаболическое тепло, поэтому его диапазон был ограничен сокращающимися более теплыми водами. Ископаемые остатки подтверждают отсутствие мегалодонов в регионах по всему миру, где температура воды значительно снизилась в течение плиоцена. Однако анализ распределения мегалодона во времени предполагает, что изменение температуры не сыграло прямой роли в его исчезновении. Его распространение в миоцене и плиоцене не коррелировало с тенденциями потепления и похолодания; в то время как численность и распространение снизились в течение плиоцена, мегалодон действительно показал способность обитать в более холодных широтах. Он был обнаружен в местах со средней температурой от 12 до 27 ° C (от 54 до 81 ° F) с общим диапазоном от 1 до 33 ° C (от 34 до 91 ° F), что указывает на то, что глобальная протяженность подходящей среды обитания не должны были сильно пострадать от произошедших изменений температуры. Это согласуется с доказательствами того, что это был мезотерм.
Изменяющаяся экосистема
Мегалодон мог стать соразмерным с более мелкими видами усатых китов, такими как Piscobalaena nana.
Морские млекопитающие достигли своего наибольшего разнообразия в миоцене, например, с усатыми китами с более чем 20 признанными миоценовыми родами по сравнению только с шестью существующими родами. Такое разнообразие представляло собой идеальную среду для поддержки суперхищника, такого как мегалодон. К концу миоцена многие виды мистицетов вымерли; выжившие виды могли быть более быстрыми пловцами и, следовательно, более неуловимой добычей. Кроме того, после закрытия Центральноамериканского морского пути разнообразие и численность тропических китов уменьшились. Исчезновение мегалодона коррелирует с упадком многих мелких ветвей мистицетов, и вполне возможно, что он полностью зависел от них как источника пищи. Кроме того, вымирание морской мегафауны в плиоцене привело к исчезновению 36% всех крупных морских видов, включая 55% морских млекопитающих, 35% морских птиц, 9% акул и 43% морских черепах. Вымирание было селективным для эндотерм и мезотерм по сравнению с пойкилотерм, подразумевая причинно-следственную связь с уменьшением количества пищи и, таким образом, соответствовало мезотермию мегалодона. Мегалодон мог быть слишком большим, чтобы поддерживать себя на истощающихся морских пищевых ресурсах. Похолодание океанов во время плиоцена могло ограничить доступ мегалодона к полярным регионам, лишив его мигрировавших туда крупных китов.
Конкуренция со стороны других хищников морских млекопитающих, таких как макохищные кашалоты. который появился в миоцене, а косатки и большие белые акулы в плиоцене, возможно, также способствовали упадку и исчезновению мегалодона. Летописи окаменелостей указывают на то, что новые китообразные-китообразные обычно обитали в высоких широтах в течение плиоцена, что указывает на то, что они могли справиться со все более преобладающими температурами холодной воды; но они также встречались в тропиках (например, Orcinus sp. в Южной Африке). Крупнейшие макрохищные кашалоты, такие как Livyatan, наиболее известны с миоцена, но сохранились до плиоцена, в то время как другие, такие как Hoplocetus и Scaldicetus, дожили до ранний плейстоцен. Они, возможно, занимали нишу, подобную той, что у косаток, прежде чем в конечном итоге быть заменены ими.
Вымирание мегалодонов подготовило почву для дальнейших изменений в морских сообществах. Средний размер тела усатых китов значительно увеличился после его исчезновения, хотя, возможно, из-за других, связанных с климатом, причин. И наоборот, увеличение размера усатых китов могло способствовать исчезновению мегалодонов, поскольку они, возможно, предпочли охотиться за более мелкими китами; Следы укусов на крупных китах могли появиться от акул-падальщиков. Мегалодон мог просто стать одновременно исчезнувшим с более мелкими видами китов, такими как Piscobalaena nana. Вымирание мегалодона оказало положительное влияние на других высших хищников того времени, таких как большая белая акула, в некоторых случаях распространившись на регионы, где мегалодон стал отсутствовать. Исследование 2019 года, посвященное зубам мегалодона из северной части Тихого океана, показало, что он вымер гораздо раньше, примерно 4–3,6 миллиона лет назад, до того, как вымерли типичные объекты добычи, как из-за изменения климата и, как следствие, фрагментации ареала, а также из-за конкуренции со стороны великих белый.
В популярной культуре
HMS Challenger обнаружил зубы мегалодона, которым было ошибочно датировано возрастом от 11000 до 24000 лет.
Мегалодон изображался в нескольких произведения художественной литературы, включая фильмы и романы, и продолжает оставаться популярной темой для художественной литературы с участием морских чудовищ. Три отдельных мегалодона, двое взрослых и один молодой, были изображены в документальном сериале BBC 2003 года Морские чудовища, где он определяется как «опасность» той эпохи. History Channel в Jurassic Fight Club изображает мегалодона, нападающего на Brygmophyseter кашалота в Японии. В нескольких фильмах изображен мегалодон, например Shark Attack 3: Megalodon и Mega Shark series (например, Mega Shark Versus Giant Octopus и Mega Shark Versus Крокозавр ). Акула появляется в видеоигре 2017 года Ark: Survival Evolved. Некоторые истории, такие как Джим Шепард Тедфорд и Мегалодон, описывают повторное открытие акулы. Стив Альтен Мэг: Роман глубокого террора изображает акулу, охотящуюся на динозавров, с ее прологом и обложкой, изображающими мегалодона, убивающего тираннозавра в море. В сиквелах книги также фигурирует мегалодон: Траншея, Мэг: Изначальные воды, Мэг: Адский аквариум, Мэг: Ночные охотники, Мэг: Поколения и Мэг: Истоки, а также есть экранизация под названием <26.>The Meg выпущен 10 августа 2018 года.
Псевдодокументальный фильм Animal Planet Mermaids: The Body Found включал встречу между стручками за 1,6 млн лет. русалок и мегалодона. Позже, в августе 2013 года, Discovery Channel открыл свой ежегодный сериал Shark Week еще одним телевизионным фильмом Megalodon: The Monster Shark Lives, противоречивым документальным фильмом о существо, которое представило предполагаемые доказательства, чтобы предположить, что мегалодон был все еще жив. Эту программу критиковали за то, что она полностью вымышленная; например, все изображенные предполагаемые ученые были оплачиваемыми актерами. В 2014 году Discovery повторно выпустил в эфир «Чудовищную акулу из жизни» вместе с новой одночасовой программой «Мегалодон: новые доказательства» и дополнительной беллетризованной программой под названием «Акула тьмы: гнев подводной лодки», что вызвало дальнейшую негативную реакцию со стороны СМИ и СМИ.
Сообщения о предположительно свежих зубах мегалодона, например, сделанные HMS Challenger в 1873 году, ошибочно датированные примерно 11000-24000 годами, вероятно, являются здоровыми зубами -сохраняется толстой минеральной коркой осадком диоксида марганца, поэтому имеет более низкую скорость разложения и сохраняет белый цвет во время окаменелости. Зубы ископаемых мегалодонов могут различаться по цвету от кремово-белого до темно-коричневого и серого, а некоторые ископаемые зубы могли быть переотложены в более молодой слой . Утверждения о том, что мегалодон может оставаться неуловимым в глубинах, подобно мегалодой акуле, обнаруженной в 1976 году, маловероятны, поскольку акула жила в теплых прибрежных водах и, вероятно, не могла выжить в холодных и бедных питательными веществами глубоководная среда.
Зубы мегалодона — это ископаемое в штате Северной Каролины.
См. Также
 Портал акул
Портал акул
- Список доисторических хрящевых рыб
- Доисторические рыбы
- Крупнейшие доисторические организмы
Примечания
Ссылки
Дополнительная литература
- Dickson, KA; Грэм, Дж. Б. (ноябрь – декабрь 2004 г.). «Эволюция и последствия эндотермии у рыб». Физиологическая и биохимическая зоология. 77 (6): 998–1018. DOI : 10.1086 / 423743. PMID 15674772.
- Kent, Bretton W. (1994). Ископаемые акулы региона Чесапикского залива. Колумбия, штат Мэриленд: Иган Рис и Бойер. ISBN 978-1-881620-01-3. OCLC 918266672.
Внешние ссылки
- Рост супер-хищных акул
- Кархарокл: вымершая акула-мегалодон
- Палеонтологические видео
- Палеонтолог Марк Ренц показывает один из самых больших зубов мегалодона, обнаруженный на YouTube
- Shark Week Специально о мегалодоне с Пэтом Маккарти и Джоном Бабиарцем на YouTube с комментариями о его исчезновении.
- Ископаемые зубы мегалодона показывают свидетельства существования питомника акул, которому 10 миллионов лет на YouTube
- Взгляд эксперта: информация о мегалодоне на YouTube (с участием эксперта Даны Эрет)
- Ламниформные акулы: 110 миллионов лет господства над океаном на YouTube (с участием эксперта Микаэля Сиверсона)
- Взлет и падение неогеновых гигантских акул на YouTube (с участием эксперта Бреттона Кента)
- Мур, Калли (19 декабря 2018 г.). «Почему мегалодон (определенно) вымер». PBS Eons — через YouTube.